« 2005年09月 | メイン | 2005年11月 »
2005年10月31日
ハロウィン
週末はハロウィン。どんなに街がにぎやかになるんだろうか、と少し期待してたのだが、僕が住んでる Fort Lee では普段と全く変わらず、少し拍子抜け。仮装してる人はおろか、かぼちゃも見かけなかった。Fort Leeは韓国人街だからハロウィンは祝わないのかな?かといって、Newark も別段、変わったところはなかった。むーん。これだったら薬作ハロウィンのほうがそれっぽかったかも。
投稿者 sfujisawa : 21:24 | コメント (0)
2005年10月28日
Phys Rev Lett 10/28
Entangled Networks, Synchronization, and Optimal Network Topology
Luca Donetti, Pablo I. Hurtado, and Miguel A. Munoz
「Entangled network」という新たなネットワークtopologyを見つけたよー、という論文。
最近はやりの、Network に関する研究(たとえば Small-world network など)は、おもに topology に焦点をあてたものが多い。確かに、人間関係のネットワークやWWWでのネットワークの研究では、topologyのみに注目してても、十分面白い。しかし、Neural networkでは、topologyに加えてdynamicsも知りたい。あるtopologyをもつネットワークにおいて、各ノード(ニューロン)が時々刻々とある活動をしているときにネットワーク全体としてどういうdynamicsになるか、ということに興味があるわけである。
ところが、BarabasiにしろWattsにしろNewmanにしろ、この手の研究で有名な人たちは、dynamicsにはあまり興味がないようです。(Barabasiなんか、scale-free network の結合行列の固有値まで求めているくせに、dynamicsには目も向けてない。。)
今回の論文は、topologyとdynamicsの両方に注目した論文です。
--
Pecoraの論文(PRL1998やPRE1998など)では、ネットワークのsynchronizationの安定性に関する議論をしています;
ひとつの振動子のdynamicsは、
dx / dt = F(x)
で記述できるとします。XはベクトルでもOKです。(Pecora論文では、x を3次元ベクトルとして Rossler Attractor などを考えたりしています。)この振動子が、N個存在して、お互いがinteractしているとき、i 番目の振動子の微分方程式を、
dxi / dt = F(xi) + σ Σj Lij H(xj)
と記述します。ここで、L は結合行列で、Lii = -k (kは、ノード i が有する結合数) 、Lij = 1 (ノード i と j が結合があるとき)、Lij = 0(ないとき)とします。(つまり、L は対称行列。)ここから、すべてのノードがsynchronizeしているときからの摂動を評価することにより、synchronizationの安定性を評価する、という流れみたいです。(途中はあんまりよく理解できなかった。)
結局、行列 L の固有値をもとめて、
固有値比 Q = (最大固有値) / (0以外の最小固有値)
が小さいほど安定性が高い、ということらしいです。トポロジーによって決まる結合行列 L から synchronization dynamics の安定性が議論できる、というのがミソ。
それでは、ネットワークのノード数 N と、平均結合数
まずはじめに、例として、small-world network や random network や linear chainなどの Q を計算している。(この中では、small-world の Q がいちばん小さいらしい。)つぎに、Q が小さくなるように、これらのネットワークから(あるアルゴリズムによって)ノードをひとつひとつつなぎ変えていく。そうしてsその極限で得られたのが、この論文のFig1の右側のようなネットワーク。このネットワークの特徴は、
・各nodeの性質が非常に似通っている。
・コネクションはとても複雑で入り組んでいる。
・Modularityは乏しい。
など。このネットワークは、“ Entangled network ” (もつれネットワーク?)と名付けられてます。
実際、神経細胞のネットワーク(たとえばCA3のリカレントネットワーク)を考えてみると、small-worldよりもこっちのentangled networkの方が近いかもね。なんとなくですけど。
まあもっとも、この論文やPecora論文で言うところのsynchronizationと、神経生理学で興味のあるsynchronizationとではまた意味が違うのですが、まあでもnetworkのdynamicsに焦点をあてている論文、ということで長々と書いてみました。
投稿者 sfujisawa : 20:21 | コメント (0)
2005年10月25日
実験
最近はずっとこまごまとした実験。今はシリコンプローブを目的部位に向けて動かしているところ。一日に少しずつしか移動させないので、いまだ到達していない。(目的部位に到達したかどうかは、記録される波形などでだいたい把握できる。)ちゃんと計算通りに到達するかどうか、ほんとにヒヤヒヤする。毎日が不安でしょうがない。
さて、慢性実験をはじめると一日の終わるのが早い。しかも単調な作業ばっかりなのであまりネタにならない。(というわけで更新頻度低下中です。)
投稿者 sfujisawa : 20:37 | コメント (0)
2005年10月22日
Local cue と Distal cue
というわけで、place cell の place field 形成における、Local cue と Distal cue の役割の違いについて、最近の論文をベースに簡単にまとめてみます。
(以下の文章では、「実験部屋」は実験を行う部屋全体の空間(つまり distal cue)、「実験箱」は実験部屋の中に置いてあり、ラットがその中で課題を行う比較的狭い空間(つまり local cue)をさします。)
(1) Knierim 2004 Nature の実験:
ある実験部屋(distal cue)とある実験箱(local cue)の組み合わせで、ラットに場所を学習させる。ここで、実験箱を実験部屋に対して反時計回りに何度か回転させる。このとき、place field が反時計回りに回転していれば、その place cell は local cue をコードしており、、place field が時計回りに回転していれば、その place cell は distal cue をコードしているのがわかる、という実験ロジック。Place field が Local cue と Distal cue のどちらに引きずられるか、ということですね。
実験結果は、
CA1では、Local 14.3% Distal 12.9%
CA3では、Local 51.5% Distal 7.7%
つまり、CA3では、かなりの place cell が local cue をコードしていることが分かる。CA1では、Local でも Distal でもない(つまり、place fieldがなくなったり、ぼやけたり、新しくできたりした) place cellがけっこう多い。
(2) Moser 2004 Science の実験:
Moser 2004 science の論文でも、Knierimの実験に近い結果が得られている。
実験箱Aから実験箱Bの変化に対して(実験部屋は同じ)、CA3は place field が overlap することが少ないが、CA1は place field が overlap することが多い、という結果。解釈としては、① CA1が distal cue で、CA3が local cue、あるいは、② CA3のほうがCA1よりも、 pattern separation 能力が高い。
(3) Moser 2005 Science の実験:
このエントリー参考。実験部屋(Distal cue)は同じで、実験箱(Local cue)を変えた場合、一見、place cellのコードする領域が変わったように見えるが、実は発火数が変わっているだけで、コードしている領域は同じだ、というないよう。(CA1&CA3野)。つまり、Local cueは発火数、すなわち"Gain"によってコードしている、という解釈。(昨日のトークで、Moserは、場所の変化に対して発火数のみ変わるものを"Rate remapping"、Place field と発火数の両方が変わるものを"Grobal remapping"と名付けて、これらの違いが生じる回路的なメカニズムについていま研究しているみたいなことを言ってました。)
(4) O'Keefe 2005 Science の実験:
O'Keefeの実験では、丸い実験箱を徐々に四角い実験箱に変えていったら、place cell の place field は、ある時点で、不連続に、丸い空間の place field から四角い空間の place field へ遷移するという結果が得られている。つまり、丸と四角の中間の空間であっても、丸い空間か四角い空間のどちらかに認識されているということですね。つまり、アトラクターが存在する、というわけですね。
ここで、実験箱を Local cue と考えると、実験は同じ実験部屋でやっているので、CA1は完全にLocal cueをコードしていることになる。
(ギューリーはことあるごとにこの論文を絶賛していますね。丸い空間と四角い空間で完璧にplace fieldが分離されてるのがすごい、と。ところで、このようにplace fieldが完全に分離されるにはどれくらい時間がかかるのでしょう?この論文のMethodをみてみると、だいたい1週間と書かれています。でも実際は、Pre-trainingをもう少し長い時間やっているかもしれないですね。O'Keefe Nature2002では、円形箱と四角型箱で完全に place field が分離するのに20日近くかかっていますからね。Moser実験とのデータの違いも、この辺のPre-training時間にあったりするのかもしれないです。)
(5) Moser 2005 Neuron の実験:
これはおととい書いたやつですね。O'Keefeと同じく実験箱を円形→四角形に変形させていったら、O'Keefeのように一気にplace fieldが遷移する細胞と、そうではなく徐々にplace fieldが変形していく細胞の両方が存在する、という結果。
昨日のトークでも話してたのですが、徐々に変形する細胞がいるからと言って、アトラクターの存在を否定するわけではない、と主張していました。というのは、このような細胞も、円形から四角形に変形していくときと、四角形から円形に変形していくときでは、place fieldの遷移の仕方が違うというデータが得られているので(Fig4(CA3)&Fig6(CA1))、つまり、「ヒステリシス(Hysteresis)」が存在する、というわけです。(ヒステリシスが存在するということは安定点の存在を示唆しますよね。)(ヒステリシスは電磁気学や固体物理学などでよく出てきますが、入力に対する応答がその履歴に依存しているということです。直感的な図(ハーマン・ハーケンの本より))
--
さて、Local cue と Distal cue が CA1細胞の place field に与える影響をまとめると、
Moser-McNaughtonの考え方では、CA1では、
Local cue は、place cell の、place field は変えずに発火数(gain)を変える。
Distal cue は、place cell の、place field そのものを変える。
O'Keefeの考え方では、CA1では、
Local、Distal に関わらず、place field が変化する。
で、Contradictoryである。(というのがギューリーの解釈)
もっとも、O'Keefeの実験で使っている実験箱は、かなり大きい(一辺 102cm、高さ77cm)。(ちなみにMoserのは、一辺 80cm、高さ50cm)。もしラットの視界に入ってくるものが実験箱の壁面だけで実験部屋が見えないなら、実験箱を local cue と考えるのはおかしいかもしれない。
そうしてみると、Local cue / Distal cue という単純な分類ではダメなのかもしれませんね。(ギューリーは、Local cue、Distal cue と言う言葉を好んで使うんですけど。)Distal cue はむしろ「Context」と言い換えた方がいいかもしれないですね。Moserの実験ではラットは実験部屋をContextとして認識してて、O'Keefeの実験では実験箱をContextとして認識しているのかもしれない。
あんまりうまくまとまりませんでしたね。またそのうち蒸し返してみます。
投稿者 sfujisawa : 20:49 | コメント (0)
2005年10月21日
Moserのトーク
ちょうどタイムリーなことに、今日の夕方、コロンビア大の Medical campus で Moser夫妻のトークがあったので、聞きに行ってきました。
May-Bitt Moser(妻)のほうが Entorhinal cortex の "grid cell" について。話のメインは、彼らの Nature 2005 でしたが、何個かNewのデータも入ってました。
ここで、簡単に "grid cell"の特徴についてまとめてみると、
(1) grid cell の place field は等間隔にgrid状に存在する。gridの間隔は30~50cmと、比較的広い。gridの交線はだいたい 60°で一定。(Nature 2005 Fig1-2)
(2) 大きい実験箱にいるときも小さい実験箱にいるときも、grid の間隔は変わらない。(Fig1)
(3) grid cellは、ECのII/III層だけではなく、V/VI層にも存在する(New)。(II/III層が海馬へのInputで、IV/Vそうが海馬からのOutputであることを考えると、入出力のどちらにもgrid cellがあることは少し不思議な感じがしますね。)
(4) 各層で、層の深い方に行くほど、grid間隔の大きい細胞が多い。(Fig2)
(5) lacal cueの位置をずらすと、gridもそれにつれられてずれる。(Fig4)
(6) 暗い環境に変えても、gridは保持される(Fig5)
(7) 新規環境でも、1分ぐらいでgrid状の place field が形成される。(Fig6)
(8) 同じgrid cellで、同じ環境であれば、gridの位置は、trialごとに変化することはなく、安定している。
(9) しかし、同じgrid cellで違う環境の場合(例えば円形の実験箱と四角形の実験箱)、gridの間隔は変わらないが、gridの位置が少し(10cmぐらい)シフトする。(New)
(10) ECには、grid cell意外にも、head directionに反応する細胞もある。II/IIIではgrid cellの方が比較的多いが、V/VI層ではhead direction cellの割合の方が多くなる。(ちなみに、head direction cellはsubiculumにも多いことが知られています。)
で、Moser妻は、このような place fieldは、① Landmark-basedに形成されるのか、あるいは② Self-motion-basedに(つまりpath integration)によって形成されるのか、ということをdiscussionしていました。上述の(2)や(6)や(7)のデータからは、path integrationかと考えられるが、(5)のデータからするとLandmark-baseなので、まだ何とも言えない、という結論でした。それにしても、やっぱり面白いですね、grid cell。海馬の入力もとであるECにこんな細胞があるとは、ほんとに驚きです。
次に、Moser旦那の方は、海馬CA1、CA3、DGやECでのplace fieldのremappingの話など。話のストーリー的には、海馬ではどこで pattern separation と pattern cmpletion が行われているか、という内容。
メインの話は、彼らの Science 2004a,2005(参考)、Neuron 2005(昨日のエントリー参考)とかでした。
面白かったのは、DGでも、Neuron 2005の円形の実験箱と四角形の実験箱への7段階変形の実験をやっていて、DGでは、2段階目か3段階目ぐらいでplace field が変化してしまう、というデータ(unpublished)。つまり、DGはpattern separationの機能が強い、とMoserは述べてました。
ところで、以前ギューリーがラボミーティングの時に話してたのですが、Moserのラボにはテクニシャンが7人もいるらしいですね。ノルウェーの研究界のホープらしくて、研究費をばんばんもらっているらしい。その話をしてるとき、うちのラボの誰だったかがギューリーに質問。「なんでうちのラボにはテクニシャンがひとりもいないんですか?」ギューリーの答え「テクニシャンが全部やってくれたら、おまえらポスドクの勉強にならないからだよ。」 うーん。。
投稿者 sfujisawa : 20:27 | コメント (0)
2005年10月20日
Neuron 10/19
Progressive Transformation of Hippocampal Neuronal Representations in ""Morphed"" Environments
J.K. Leutgeb, S. Leutgeb, A. Treves, R. Meyer, C.A. Barnes, B.L. McNaughton, M.-B. Moser, and E.I. Moser
Moser、またかぃ!
ラット海馬CA1とCA3での place cell についての論文。実験のパラダイムは、前回のO'Keefeの論文(Wills et al Science 2005)(参考)とほぼ同じ。円形の実験箱と四角形の実験箱を使う。Place cell は円形箱と四角形箱では、place field が異なる(そうなるまでプレトレーニングで学習させる)。このとき、実験箱を、円形から四角形に徐々に(7段階ぐらいで)変形していったら、place cellはどう発火するか、という実験。
O'Keefeの実験では、丸い空間を徐々に四角い空間に変えていったら、place cell の place field は、ある時点で、不連続に、丸い空間の place field から四角い空間の place field へ遷移するという結果が得られている。今回のMoser論文では、O'Keefe論文のようにplace fieldが不連続に遷移するという細胞(Figure3、"Sigmoid")と、連続的に段階的に変化するという細胞("Linear")の両方が存在したという結果(CA1、CA3ともに)。
CA1とCA3での違いについては、CA3の place cell ほうが、CA1のよりも、異なった環境(円形箱と四角形箱)をより分離する傾向がある、ということらしい。(Place field の overlap率のデータなどから。)
さて、最近似たような論文がでてこんがらがってきたので、明日ぐらいまたちょっとまとめてみます。
投稿者 sfujisawa : 22:25 | コメント (0)
2005年10月19日
オペ
今日はオペをした。シリコンプローブ2本刺し。オペは9時間ほどかかった。やれやれ。でもまあ、見た目はなんとかうまくいった感じ。しかし、実際に目的部位にきちんと到達してきちんと記録がとれるかどうかはもう何日かしてみないと分からない。不安な日々が続く。
投稿者 sfujisawa : 22:26 | コメント (0)
2005年10月18日
準備など
Neuroscience学会まであと一ヶ月弱ほど。学会までにひとつ慢性実験を終わらせたいので、明日オペを予定している。今日はその準備など。しかし、どうもみんな考えることは同じみたいで、先週・今週と手術室も工作室もえらい混雑である。なかなか作業が進まんね。
それにしても、オペが成功するか失敗するかは結構でかいので、オペの日が近づくとホント緊張してくる。なんかそわそわしてくる。ネガティブな気分におそわれる。なんとなく試験前のような気分。
投稿者 sfujisawa : 22:33 | コメント (0)
2005年10月17日
散髪
先月、日本に一時帰国したときに隆二に会ったのだが、彼は相変わらず坊主頭だった。バリカンを使って自分でカットしているらしい。手軽でいいよ、やってみたらー、と勧められた。
なんとなく羨ましくなって、日本にいる間に、秋葉原のヨドバシカメラで、ナショナルのセルフヘアカッターというのを買ってしまった。(そのままアメリカに持ってきた。)
で、最近髪が伸びてきたので、さっそく昨日、それを使って刈ってみました。
結論を言うと、なかなか、取扱説明書にモデルででているお兄ちゃんのようにきれいには刈れないですね。側面にムラができて、むしろコボちゃんに近いかも。しかも、僕は手鏡を持っていないので、後ろがどうなっているのか確認できない。
まあ、でも、手軽なのは確かにいいです。興味ある人はトライしてみてください。
投稿者 sfujisawa : 22:33 | コメント (0)
2005年10月14日
PCAとSVD
ブザキラボと、隣のケン・ハリスラボ(ケンはブザキ研出身なのでラボどうし仲がよい)は、数物系出身の理論家が多い。全体の1/3ぐらいが理論家かな。解析系のプログラムなどは、理論系の人がバリバリ書いてくれる。実験系の人は、プログラムの使い方は分かるが、しかし原理まではなかなか勉強できない。それで、ハリスラボのポスドクのカリーナ(彼女は数学出身)が実験系の人のために、解析用の数学を簡単に説明してくれる勉強会を毎週金曜にしてくれているらしい。その勉強会の存在を先日知ったので、今回から僕も参加させてもらうことにする。
今日のお話は、PCA(principal component analysis:主成分分析)と SVD(Singular Value Decomposition:特異値分解)の違いについて。カリーナが講義室で黒板を使って30分ぐらいで説明してくれる。
さすが数学出身だけあって、話がシンプルですごいわかりやすかった。内容を全部書きたいところだけど、webで数式を書くのは大変なので、省略してまとめてみます。(mimetexとかを入れればいいんだろうけど、入れ方が分からない。。)
N次元のデータが、n個あったとする(ただし、n >= N )。すなわち、
y1 = (y11, y12,・・・, y1N )
y2 = (y21, y22,・・・, y2N )
・・・
yn = (yn1, yn2,・・・, ynN )
この、n行N列のデータ行列を A とおくとき、A の共分散行列は、
AT A
で求められる(詳細は省略)。PCAは、この AT A の固有値問題に帰着される。
で、SVDは、行列 A に対して、 結局 AT A の固有値問題に帰着されるらしいので、要するに、PCAとSVDは同じことらしい。
AT A は当然 N×N の対象行列になるので、N個の非負固有値が存在するはずで、固有値問題が解けるわけですね。
ここまで分かれば、matlabで簡単にPCA解析できますね。普通に固有値・固有ベクトルを求める eig 関数を使ってもいいし、svdという関数があるのでそれを使えばもっと簡単ですね。
例えば、Aをn行N列のデータ行列とするとき、
[U,S,V] = svd(A);
と入力すると、A=U S VT となるよう特異値分解してくれて、これはPCAを行っているのと等価なわけですね。(なんと一行でPCAができてしまう!)ここで、V は主成分結合係数ベクトルを並べた行列。データ行列を主成分軸の張る空間に移したければ、A*V(あるいは U*S)で計算できますね。
(↑これで正しい?)参考ページ。
投稿者 sfujisawa : 22:52 | コメント (0)
Science 10/14
Observing Others: Multiple Action Representation in the Frontal Lobe
Koen Nelissen, Giuseppe Luppino, Wim Vanduffel, Giacomo Rizzolatti, and Guy A Orban
サル、fMRI。ミラーニューロンネタ。(勉強しなければ。。)
投稿者 sfujisawa : 22:51 | コメント (0)
2005年10月13日
脳波メモ
前回のラボセミナーはアントンのJournal clubで、Tononiのこの論文を紹介したのだが、脳波の話になると、俄然ギューリーが興奮してくる。とくにこの論文では、Fig2の覚醒下でのTMS-responseを見て、「これは単なるalpha波のリセットじゃねーのか」と声高に主張していた。(Fig1でalpha波がきれいに見えている(つまり覚醒下で目をつぶっている状態)のが理由。)まあ、それはともかくとして、ギューリーは脳波(やLFP)の話になると話が止まらなくなる。どうやら重度の「脳波マニア」みたい。話を聞いてるだけで勉強になって面白いのですが、話がマニアックになるとついていけなくなってしまうので、少し僕も勉強することにしました。
今回は、NonREM睡眠時でのCortexで見える脳波についてまとめてみました。
(1) Slow oscillation: <1 Hz, generally 0.5~1 Hz
発生源:Neocortex。
理由(i) thalamusを切除してもcortexでslow oscillationが残る。[Ref]
理由(ii) cortexを切除すると、thalamusではslow oscillationが消える。[Ref]
(いわゆるUP-DOWN stateは基本的には麻酔のartifactと考えられていますが、脳波的にはslow oscillationに対応すると考えればいいと思います。)
(2) Spindle: 12~15Hz (Human), 7~14 Hz(Cat), Rat ~14 Hz(Rat)
発生源:thalamic reticular (RE) neuronがペースメーカー 。[Ref]
ただし、広域でのspindleの同期には、Neocortexが必要。[Ref]
(3) Delta: 1~4Hz
発生源:thalamus起源(thalamocortical neuron)とneocortex起源の2種類ある。
端的な模式図がありました。
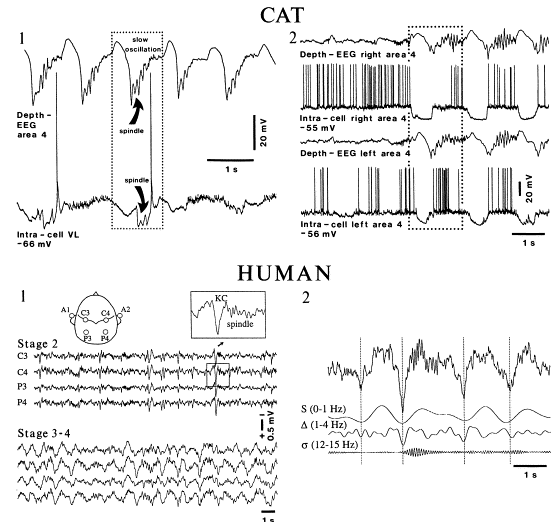
Steriade の Neurosci 2000のFigure8をぱくりました。(Steriade先生、すいません。。)
上図はCatのintra/extra記録と、humanのEEGを比較したもの。
Slow oscillationは、NonREM中で、intra記録でUPからDOWNに遷移するときに、extra記録で上向きに観察されている波。この図では、Slow oscillationの後にspindleが続いている。(この、slow oscillation-spindleのペアは、よく観察される。)
Human(scarp)でのEEG記録では、slow oscillationはNREM時に下向きの(この記録では下向きが+らしい)大きな波として観察される。(これを、別名K-complex(KC)というらしい。)NREMのStage2ではslow waveにspindleが続くのが観察され(KC/spindle)、NREMのState3~4ではslow waveにdeltaが続くのが観察される(KC/delta)。(脳波による睡眠 stage の分類(ヒト)の一覧表は、このページにありました。)
Steriadeによる、slow wave (K-complex)、spindle、delta波の生成メカニズムの模式図。
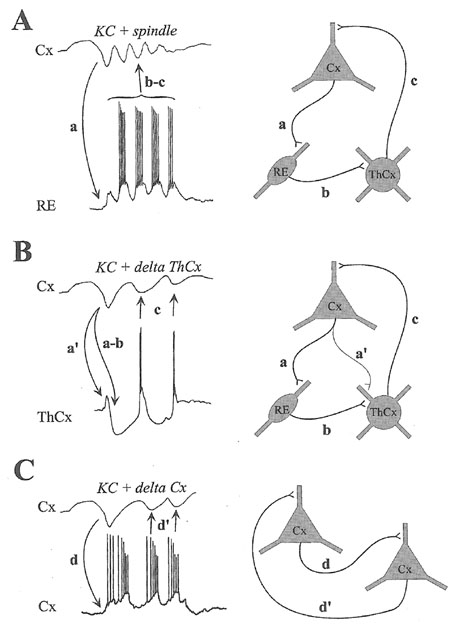
Steriadeの本のFig3.43をぱくりました。
ところで、Slow oscillationが初めて記述されたのは、1993年と結構新しい。それ以前までは、Slow wave(徐波、4Hz以下の波)として一緒にされていた。Steriadeの1993年の論文(ネコのintra記録でslow oscillationを発見し、ヒトEEG記録で確認)により、slow oscillation (<1Hz)とdelta (1-4Hz)がはっきりと分けられるようになる。
さて、ほんとは、Cortex/Hippocampus間やCortex/Cortex間での脳波の同期・伝播や、脳波リセットなどについて(とくに覚醒下と睡眠下での違いなど)を書きたかったのですが、今日は疲れたのでまた今度まとめてみます。
*以上のまとめは以下の本の3.2節を参考にしました↓。Steriadeも重度の脳波マニア。
Neuronal Substrates of Sleep and Epilepsy、Mircea Steriade (Cambridege Press, 2002) サンプル
(ところで、Amazonでこの本のカスタマーレビューを書いているbrainfreakさんって、なんとなくpooneilさんのような気がするのですが。。気のせい?)
投稿者 sfujisawa : 22:06 | コメント (6)
2005年10月12日
鼻水
先週末からずっと雨続きで、最近は気温が10度前後まで低下。おかげで、鼻水とくしゃみが止まらなくて困る。別に風邪と言うほどでもないので別にいいのだが、ただラットのトレイニングの時に困る。どうも僕のラット君は音に敏感で、トレイニング中に僕が鼻をズズッとすすると、ラット君はビクッとしてキョトンとした顔をされてしまう。くしゃみなどしようものなら、ビヨンと飛び跳ねてびっくりする。吉本新喜劇か、っていうぐらいリアクションでかい。仕方ないのでトレイニング中は鼻水もくしゃみも我慢せざるを得なくて、けっこうくるしい。鼻炎スプレーが必要かもね。
投稿者 sfujisawa : 21:26 | コメント (2)
Nature 10/12
メモ。
Repeated cocaine exposure in vivo facilitates LTP induction in midbrain dopamine neurons
Qing-song Liu, Lu Puand Mu-ming Poo
Pooは最近こんなことやってんですね。コカイン投与はin vivo(腹腔)ですが、電気生理はスライスみたいです。
投稿者 sfujisawa : 21:24 | コメント (0)
2005年10月11日
エディター来訪
先日、ギューリーから、「Important」というタイトルのメールが送られてくる。「11日に Nature Neuroscience の Editorのナラシムハン博士がうちのラボに訪問に来るので、印象を良くするために当日はみんなactiveに実験しててね。」お客さん来るときにがんばるのはどこのラボも同じらしい。。
さて、NN のEditorとはどんな人かと思いきや、けっこう若いのでびっくりした。どうも、Nature系は専任の職業 Editor が Editional workをするみたいですね。
夕方、30分間ほど、ナラシムハン博士が NN の Editional workについてトーク。NNは月に170報ほどの投稿があり、そのうち70%が一発reject、最終的な採択率は10%程度とか。あとは選考基準や(Reject後の) Appeal letterを書くときの注意点などの話。笑えたのが、「こんなAppeal letterを受け取ったことがあります」と言って見せてくれたletterで、ナラシムハン博士宛で「I'm afraid you are nuts.」と一行だけ書かれたletter(署名入り)。気持ちは分かるけどね。。
投稿者 sfujisawa : 22:08 | コメント (0)
2005年10月10日
週末
週末は、マンハッタンでテロ予告の情報(けんじさんに教えてもらった)があったので、しかも土日はずっと雨だったため、家にずっと滞在。家事と読書にいそしんだ週末でした。気温もこの雨でぐっと下がってきた模様。さすがに半袖では寒くなってきたので、段ボールから長袖を引っ張り出してきたり。ところで、マンハッタンは特に何事もなくまあよかったわけですが、無かったら無かったでまたいろいろ問題があるらしい。
投稿者 sfujisawa : 20:46 | コメント (0)
2005年10月07日
Science 10/7
ところで、来年のSfNの annual meeting は、アトランタに変更になったみたいですね。
Failure to Detect Mismatches Between Intention and Outcome in a Simple Decision Task
Petter Johansson, Lars Hall, Sverker Sikström, and Andreas Olsson
以下、先週のScience。読むの忘れてた。
Breakdown of Cortical Effective Connectivity During Sleep
Marcello Massimini, Fabio Ferrarelli, Reto Huber, Steve K. Esser, Harpreet Singh, and Giulio Tononi
ヒト、TMS(経頭蓋的磁気刺激法)とEEG(60電極)。覚醒時とNonREM睡眠時で、皮質にTMS刺激を与えたときに、TMS-evoked response をEEGで調べて比較した、という内容。覚醒時では、TMS-evoked response は 300ms ほど続き、かつ数センチ離れた皮質まで伝播する。しかし、NREM時では、TMS-evoked responseのamplitudeは覚醒時より大きいものの、120msほどで減衰し、また伝播も観察されなかった。このような覚醒時と睡眠時における皮質の「effective connectivity」の違いを、consciousnessと絡めて議論しています。
Direct Evidence for a Parietal-Frontal Pathway Subserving Spatial Awareness in Humans
Michel Thiebaut de Schotten, Marika Urbanski, Hugues Duffau, Emmanuelle Volle, Richard Levy, Bruno Dubois, and Paolo Bartolomeo
ヒト、Intraoperative electrical stimulationという手法。これは、Glioma(神経膠腫)患者の腫瘍摘出手術中に、摘出部位の境界を決定するために行うものらしい。頭蓋を開けた状態で、脳表の摘出予定部位に直接電極をあて、電気刺激をしてその部位の活動を一時的に抑制する。そのとき、患者が一時的に重要な障害を生じれば(たとえば会話ができなくなるとか)、その部位は摘出するのを止める、というような感じで。
ここで、(右半球の)caudal superior temporal gyrus (cSTG) および supramarginal gyrus (SMG)を電気刺激したとき、線分を二等分する課題で、障害を生じた(分離点が右にずれた)、という結果。(なので、この部位の摘出は止めた。)この部位は、Parietal-Frontal pathwayに相当する部位らしいが、このpathwayが視覚のsynmetrical processingに必要である、という結論。
この論文では特に半側性空間無視などと絡めて議論しているが、この辺は勉強不足なので、pooneilさんのサイトを復習して勉強中です。
投稿者 sfujisawa : 21:52 | コメント (0)
2005年10月06日
ラット祭り?
今日朝大学に着くと、巨大ラットのバルーンが出現していた。近くで何かプラカード持ったヒトがたくさんいたから、何かのキャンペーンだったのかな?でも、プラカードの字が小さすぎて、何のキャンペーンなのかは分からなかった。なんか効率の悪いキャンペーンだ。

話は変わりますが、CNNのあほネタのページ。
投稿者 sfujisawa : 20:55 | コメント (0)
Neuron 10/5
メモ。
Perceptual Organization of Tone Sequences in the Auditory Cortex of Awake Macaques
Christophe Micheyl, Biao Tian, Robert P. Carlyon, and Josef P. Rauschecker
サルの電気生理&ヒト。A、Bの別の周波数をもつ音を、A-B-A A-B-A ・・・と繰り返し発生させるとする。AとBの周波数の差ΔFが小さければ、「A-B-A」と聞こえるし、ΔFが大きければ、「A-A-A-A」「B-B」と別の2つのsteamとして聞こえる。この現象に対して、ヒトでの心理物理学的実験と、サル一次聴覚野でのユニット記録の実験を行ったもの。
Updating Visual Space during Motion in Depth
Nuo Li and Dora E. Angelaki
投稿者 sfujisawa : 20:54 | コメント (0)
2005年10月05日
JNS 10/5
Scopolamine Reduces Persistent Activity Related to Long-Term Encoding in the Parahippocampal Gyrus during Delayed Matching in Humans
Karin Schon, Alireza Atri, Michael E. Hasselmo, Marisa D. Tricarico, Matthew L. LoPresti, and Chantal E. Stern
ヒト、fMRI。(1)視覚のdelayed matching-to-sample (DMS) taskを行っているとき、delayの時のparahippocampusの活動をfMRIで測定。(これを、persistent activityと解釈している。)スコポラミン(ムスカリンR-アンタゴ)を投与された被験者では、このpersistent activityは減少する。(2)DMSの20分後に、DMSで使った画像を覚えているかのtask(これをlong-term momoryとしている)。スコポラミン群ではこのスコアも減少していた。つまり、DMSでのdelay時のpersistent activityと長期記憶の関係(相関)と、そのときのコリン系の役割を調べた研究。
NMDA/AMPA Ratio Impacts State Transitions and Entrainment to Oscillations in a Computational Model of the Nucleus Accumbens Medium Spiny Projection Neuron
John A. Wolf, Jason T. Moyer, Maciej T. Lazarewicz, Diego Contreras, Marianne Benoit-Marand, Patricio O'Donnell, and Leif H. Finkel
コンピューターモデル実験。ここでのstate transitionは、UP/DOWNでのDOWN←→UPのtransitionのことでした。
Behavioral Evidence That Segregation and Representation Are Dissociable Hippocampal Functions
Stepan Kubik and Andre A. Fenton
ラット、行動課題。あまりよくわからなかった。
Hormonal Cycle Modulates Arousal Circuitry in Women Using Functional Magnetic Resonance Imaging
Jill M. Goldstein, Matthew Jerram, Russell Poldrack, Todd Ahern, David N. Kennedy, Larry J. Seidman, and Nikos Makris
投稿者 sfujisawa : 20:42 | コメント (0)
2005年10月04日
Nature Neurosci 10月号
時差ぼけで10時睡眠-4時起床の生活(ある意味健康的)。げりもまだなおらん。
NNの最新号を読み忘れていたので、軽く目を通してメモ。
Dynamics of excitation and inhibition underlying stimulus selectivity in rat somatosensory cortex
W Bryan Wilent & Diego Contreras
ラットの in vivo イントラ記録。Somatosonsory cortex の thalamoreceipient neuron から。このニューロンでは、ひげを動かす方向に対して方向選択性があるらしい。その方向選択性のメカニズムについて、興奮性入力と抑制性入力それそれのコンダクタンスを詳細に測定することで明らかにしようとした論文。
Asynchronous GABA release generates long-lasting inhibition at a hippocampal interneuron principal neuron synapse
Stefan Hefft & Peter Jonas
海馬 DG で、interneuron と granule cell をダブルパッチして、interneuronの電気生理学的性質を調べたもの。この論文では、CCK-positiveなinterneuronでは、強い発火が生じた後では、somaの発火タイミングに依存しないGABAの放出(←つまりminiatureみたいなもの?)が存在する、という発見。これを、asynchronous GABA releaseと呼んでいる。PV-positiveなinterneuronではこの現象は無いらしい。
Cortical responses to invisible objects in the human dorsal and ventral pathway
Fang Fang & Sheng He
ヒト、fMRI。左右の目に別々の画像を見せられるようにした状態で(つまり視野闘争が生じている状態で)、dominantな方の目にはノイズ画像を見せて、non-dominantの方の目にはobject画像を見せる。(つまり、意識下ではobject画像は認識されていない。)このとき、dorsal pathwayとventral pathwayでどう反応が異なるか、を調べたもの。結果は、dorsalでは、このようなときも、objectが認識されていると時と同様に強く反応するというもの。しかも、object画像が、人間の顔の画像の時よりも、道具の画像の時の方が強く反応するとか。(これは、dorsal pathway(つまり"where" pathway)が、reachingやgraspingに重要な役割を持っているからではないかと考察している。)
Tuning for spectro-temporal modulations as a mechanism for auditory discrimination of natural sounds
Sarah M N Woolley, Thane E Fremouw, Anne Hsu & Frederic E Theunissen
鳥(zebra finch)、ユニット記録。(鳥はちょっと苦手。)
Timing of the brain events underlying access to consciousness during the attentional blink
Claire Sergent, Sylvain Baillet & Stanislas Dehaene
投稿者 sfujisawa : 20:56 | コメント (0)
2005年10月03日
食あたり?
日本で生ものを食い過ぎたせいか、どうやら腹を下した様子。飛行機に乗って帰米し始めるあたりから腹の調子は下り坂。セイロガンを飲みまくってみるも、いっこうに効かず。週末は腹痛と時差ぼけがピークになって、一日中ベッドに倒れ込んでいる始末であった。今日はなんとか起きあがって大学には行ったが、未だに腹痛てー。読むべき論文もたまってきているのだが、未だ本調子ならず。。むーん。

{kind=link}
{kind=link}