2006年05月20日
Neuron 5/18
Visualizing the Cortical Representation of Whisker Touch: Voltage-Sensitive Dye Imaging in Freely Moving Mice
Isabelle Ferezou, Sonia Bolea, and Carl C.H. Petersen
Free-moving rat で、 Somatosensory cortex から Voltage sensitive dye を使ったイメージング。これすごいなぁ。Movie 見た感じでもあんまりぶれてなさそうだし。
解像度は光ファイバーの口径で決まるのかな?ここでは 8μm 系のファイバーを 6×6 毎にの bundle にして、最終的に 300 × 300 の bundle にしてるみたい。
うーん、なんかそのうち Free-moving で Ca++ イメージングとかできたりするようになるのかなぁ。これは Free moving 電気生理屋からすると脅威やね。
投稿者 sfujisawa : 00:08 | コメント (1) | トラックバック
2006年05月05日
Science 5/5
Conjunctive Representation of Position, Direction, and Velocity in Entorhinal Cortex
Francesca Sargolini, Marianne Fyhn, Torkel Hafting, Bruce L. McNaughton, Menno P. Witter, May-Britt Moser, and Edvard I. Moser
Moser ラボ、Grid cell ネタ。Entrhinal cortex の各 layer (II~V) の principal neuron の特徴を調べた論文。このエントリも参考。
EC において、以下の 3 種の細胞が記録されたらしい:
(1) grid cell (Fig 2)
ある一定間隔、一定方向(60°)に place field をもつ細胞 (Nature 2005)。「gridness」の評価は、空間における発火率の auto-correlation をとって、60°& 120°がピークとなっているか、および 30°& 90°& 150°がトラフとなっているかを、計算したらしい。
Layer II に多い。
(2) head-direction cell (Fig 3)
ラットの頭の向きに反応する細胞。「directional tuning」の評価は、ラットの頭の向きと細胞の発火率の関係を計算したらしい。(統計法は Watson's U-test というのを使ったらしい)。
Layer III~V に多い。
(3) cojunctive (grid × head direction) cell (Fig 4)
Grid cell × head direction cell の特性を持つ細胞。 つまり、gridness score も高く、directianl tuning 性も高い。
Layer III と V に多い。
----
あと面白いのは、Postrhinal cortex との境界からの距離が離れるにつれて、Grid cell の Grid の間隔が大きくなる傾向がある(すべての Layer で)。(Suppl Fig 3)
投稿者 sfujisawa : 15:07 | コメント (2) | トラックバック
2006年05月01日
PLoS 4月号
Offline Persistence of Memory-Related Cerebral Activity during Active Wakefulness
Philippe Peigneux, Pierre Orban, Evelyne Balteau, Christian Degueldre, Andre Luxen, Steven Laureys, Pierre Maquet
Maquet ラボ。ヒト、fMRI。睡眠前の覚醒下の状態での、off-line processing について。
Experimental design は Fig 1 にあります。fMRI (Session I) → training → fMRI(II) → 休憩 → fMRI(III) → retraining → fMRI(IV)。課題は、spatial navigation task か、Serial reaction time (SRT) task (Sequence を覚える課題) のどちらかを行います。fMRI 中は、auditory oddball task(ある音程の音が繰り返し聞かされ、たまに違う音程の音が混じる。その違う音程の出現回数を数える課題)を行います。
Spatial navigation task 直後の session II では hippocampus に高い活動がみられ、休憩後の session III では、II に比べて parahippocampas に高い活動が見られた、という結果。また、SRT task 後では小脳に高い活動が見られた、という結果。
---
この前の Wilson の論文(覚醒下 ripple でのスパイク列圧縮現象)を読んで以来、睡眠前の覚醒下での off-line processing に興味が出てきました。
睡眠前の reactivation の研究としては、non-human では McNaughton/Wilson が昔からいろいろやっているみたいですね。Science 2002 (macaque) とか JNS 1999 (rat) とか。またそのうちまとめてみます。
投稿者 sfujisawa : 19:34 | コメント (0) | トラックバック
2006年04月24日
Nature 4/19
Weak pairwise correlations imply strongly correlated network states in a neural population
Elad Schneidman, Michael J. Berry, II, Ronen Segevand William Bialek
Retina の多細胞発火活動の統計物理的解析。pooneil さん、shuzo さんのサイトにもコメントあり。
むずかしくて、きちんと理解できたか自信がないのですが、とりあえずまとめてみます。
---
10 個のニューロンのネットワークの活動について知りたいとき、10 個のニューロンすべてに電極をさして同時記録すると、完全な情報が得られる。でも、10 個同時パッチは大変なので、2 個同時パッチをすべての組み合わせ(= 10C2 =45通)について行ったとしたら、どうだろう?直感的に考えると、いくらすべての組み合わせで 2 個パッチしたとしても、10 個同時記録をしなければ見えないような情報をかなり落としてしまっているような気がする。しかし、この論文では、10C2 回の 2 個同時記録で得られる情報量 I2 は、10 個同時記録で得られる情報量 I10 の約 90 % をカバーしている(I2/I10 ~ 0.9)ことを示している。
---
この論文のメインの主張は、大きく2つあり、
(1) 実際の retina から得られた情報パターンの分布は、最大エントロピー法より算出された分布(いわゆるボルツマン分布)とよく一致する。
(2) 最大エントロピー法による分布を考えるとき、2 つの細胞間の correlation を拘束条件として考慮したモデルが十分良い近似となっている。(3つ以上の correlation をわざわざ考えなくてもよい。)
ということです。
(1) は、統計物理では古くから知られている事実です。神経科学でも、よく応用されています。しかし、いままでは統計物からの天下りでボルツマン分布が成り立つことを仮定してから使ってたのですが、この論文では、retina ラスタープロットのパターン出現確率分布と、ボルツマン分布がよく一致していることを、実際に実験データから示しています。
(2) は、前段落で述べたような話です。
(注)この論文でいう「パターン」は、時間方向の発火パターンのことではなく、ある時間 bin でどの細胞が発火したか、という細胞番号方向のパターンです。(後述します。)
---
この論文のミソは、① Fig 1f & 2a と ② Fig 2c だと思うですが、これについて書いてみます。
実験データの解析の方法ですが、まずラスタープロットを、ある時間Δτ(ここではΔτ= 20 ms)で区切って、スパイクがあれば "1"、なければ "0" と、バイナリ化します。N = 10 個の細胞のラスタープロットを使うと、各時間 bin のデータは 10 文字の数列で表されます(Fig1b)。
fig 1f では、10 個のニューロンのラスターにおいて、ある 10 文字の数列パターンが出現する確率 (observed pattern rate (1/s): 横軸)と、もし各細胞が独立に発火すると仮定したときに計算されるそのパターンの出現確率(Approximated pattern rate: 縦軸)の関係を示しています。
たとえば、「1011001010」というパターン(cell 1,3,4,7,9 が同時に発火)を考えてみます。cell 1 の発火確率(実測値)を p1、cell 2 の発火確率を p2、・・・とします。「1011001010」のパターンが出現する確率は、p1×(1-p2)×p3×・・・(1-p10)となります。グラフから読み取ると、計算した値は約 10-8 1/s ですが、実際に観察されたのは 10-1 と 10-2 の間ぐらいです。
つまり、このパターンは実際に観測された出現率は、計算よりもはるかに高いことが分かります。
この、各細胞が独立に発火すると仮定したときに計算されるそのパターンの出現確率 inpendent model P1 は、
P1 = Πi pi (文献27の式4)
ここからが本題ですが、それでは、cell 1 の発火確率 は p1、・・・(上と同じ)で、かつ、cell 1 と cell 2 が同時に発火する確率 q12、・・・というように、2 つの細胞 i, j が同時に発火する確率が拘束条件として入っている場合、パターンの出現確率はどう計算されるか。
これは、最大エントロピー法によって求められます。結果は、
P2 = (1/Z) exp [Σhiσi +(1/2)ΣJijσiσj ] (論文の式1)
これは、いわゆる、ボルツマン分布です(※1)。これを、2次の maximum entropy distribution P2 と名付けています。
これを使って計算したパターンの出現確率 P2 と実際のパターン出現確率の関係が、Fig 2a です。(Fig 2a の青点は、Fig 1f の青点と同じです。)赤点。P1の場合と比べて、 observed と P2 がよく一致しています(←黒線上に近い位置にある)。
つまり、2 つの細胞間の関係を拘束条件として最大エントロピー法をもちいて計算されたパターン出現確率は、実験で得られたパターン出現確率をよりよく再現している、ということです。
これが、主張 (1) に相当します。
さてそれでは、cell 1,2,3 が同時に発火する確率が r123、・・・、というふうに、より高次(k 次)の同時発火確率も拘束条件として含めたらどうか?それも最大エントロピー法をつかって計算できて、すなわち k 次の maximum entropy distribution Pk です。( k = 1,2,・・,N)
高次になればなるほど、より正確に実験で得られたパターン出現確率をよりよく再現できると考えられます。
そうすると、次数が上がっていくにしたがって、どれほど正確になっていくかを知りたくなります。
そこで、k 次の maximum entropy distribution Pk が、P1 に比べてどれほど情報が増えているかを評価するために、k 次の multi-information というものを定義します。これは、(高次の)mutual information です。詳しくは文献27にあります。2次の multi-information I2 は、いわゆる mutual information です。
それで、I2/IN をプロットしたのが、fig 2a です。I2/I10 ~ 0.9 というデータが得られており、つまり 2 次の貢献度がすごく大きい、ということです。
以上が、主張 (2) に相当します。
------
(※1)
これは、Ising モデル(や Hopfield モデル)と同じです。
Ising model(Hopfield model)ではハミルトニアン H は
H = -Σ hiσi - (1/2)Σ Jijσiσj
ですが、このとき確率分布はボルツマン分布に従います。
P = (1/Z) exp (-βH)
ここで、σi は、i 番目の細胞の発火状態で、発火なら 1、非発火なら -1 です(固体物理ではスピンの向きに相当します。) Jij は、細胞 i と j の相互作用の大きさです。Z は分配関数で、βは逆温度。(この論文では温度は考慮していないので、βは出てきません。)
さて、このハミルトニアンを計算するのに、彼らは、 hi は <σi >(cell i の bin 化されたスパイクの平均発火率(出現率))、 Jij は、<σi, σj > (cell i,j の同時発火確率)の実測値を使っています。
( hi や Jij の実際の数値については、Fig 3 以降に詳しく書かれています。)
---
要するに、この論文の主張は、1 個細胞の平均発火率を hi、 2 個ペアの correlation を Jij としてハミルトニアン H を計算したときに、パターンの出現確率は、
P2 = (1/Z) exp(-H)
でよく近似できている、ということです。
結局、固体統計物理、情報統計物理の世界で発見され洗練されてきた考え方が実験神経科学でも通用するよ、ということを示した論文なわけで、面白いのは面白いのですが、本当にこの論文が nature article に相当するほどオリジナルなのかどうかは僕にはぴんときませんでした(今のところ)。
投稿者 sfujisawa : 09:43 | コメント (4) | トラックバック
2006年04月18日
Nature AOP
今日のラボミーティングの文献紹介で使われたネタ2報。
Modulation of intracortical synaptic potentials by presynaptic somatic membrane potential
Yousheng Shu, Andrea Hasenstaub, Alvaro Duque, Yuguo Yu and David A. McCormick
McCormick ラボ。実験系は ferret の cortex のスライス。Presynaptic 側のニューロンの膜電位によって、post 側の synaptic potential (EPSP) の大きさが変化する、という論文。Pre の膜電位が浅いと、EPSP は大きくなるらしい。発想の転換だね。すごいシンプル。なんでいままで誰もやってなかったのだろう。。
メカニズムは、membrane potential が浅いほど、axon での action potential の area が大きくなる(amplitude は減少するが duration が大きくなる)、というのが原因ということらしい。(Soma と axon のダブルパッチより)。
もっとも、post側が UP だと EPSP が小さくなる(Sakmann の論文とか)ので、Pre と Post で UP-DOWN が同期してたら帳消しになるじゃん。(普通は同期してる。)なので、これが生理状態でどこまで本質的かはまだ分からない。
Combined analog and action potential coding in hippocampal mossy fibers.
Alle H & Geiger JR
こちらは海馬スライス。Mossy fiber の presynaptic terminal (mossy fiber bouton, MFB) のパッチ。
Dentate gyrus に刺激を与えたとき、Soma での membrane potential の変動が、action potential とは independent に、axon を通って passive propagation によって mossy fiber bouton まで伝達されている、という話。(これを excitatory pre-synaptic potential (EPreSP) と名付けている。)
(Menbrane potential fluctuation の axon への伝達は、上述の McCormic 論文でもやっている(Fig 4)。減衰の空間定数は、McCormic 論文では λ= 417 μm、Geiger 論文では λ= 430 μmで、ほぼ同じ。)
それで、mossy fiber bouton の膜電位が高いときは、同じ action potential でも、post-synaptic potential は大きくなるらしい。
投稿者 sfujisawa : 17:42 | コメント (1) | トラックバック
2006年04月17日
JNS 4/12
Essential Role of the Hippocampal Formation in Rapid Learning of Higher-Order Sequential Associations
Ceren Ergorul and Howard Eichenbaum
Eichenbaum ラボ。Rat で high-order の Serial reaction time 課題を開発したよ、という話。
---
Serial reaction time (SRT) 課題とは、ある sequence を覚える課題を行い(たとえば)、学習が行われたかどうかは reaction time で評価する、という課題。
この課題には、first-oeder と scond-order があり、first-order conditional (FOC) sequence は、1-2-3-1-2-3-・・のように、ある item の次にくる item が一通りに決まってしまう sequence のこと。一方、second-order conditioning (SOC) sequence は、1-2-3-1-3-2-・・のように、複合的な要素からなる。
それで、Amnesia の患者では、FOC は学習できる(reaction time が減少する)が、SOC はできないらしい (Curran 1997)。また、fMRI の研究によると、SOC では medial temporal lobe の活動が高くなるらしい (Schendan et al 2003)。それで、このような high-order sequence の学習には海馬-側頭葉系が必要なのではないかと考えられてきたようだ。
そこで、Eichenbaum は その SOC task をラットで構築して調べたよ、という話。
海馬破壊ラットと control ラットとの比較実験。
実験箱には nosepoke port(鼻をつっこむ穴)が 6 つある。正しい nosepoke port 内では reward として水が与えられる。どの nosepoke port に入ればいいかは、 port の上にある LED によって cue されている。それで、sequence のタイプは、FOC3、FOC6、SOC の 3 種類ある。
・3-item first-order conditional sequence (FOC3)
6 つある nosepoke のうち、3 つの sequence が繰り返される。(例)5-4-6-5-4-6。
・6-item FOC (FOC6)
6 つの nosepoke をすべて使う。(例) 4-2-3-5-7-6。
・second-order conditional secence (SOC)
(例) 4-5-6-4-6-5。
それぞれ、correct average reaction time (cART) と Error 数を評価。
結果はやや微妙。海馬破壊群(H-group)でも、control 群でも、SOC・FOC6・FOC3 のすべてで学習は緩やかに進んでいく(cART、Error が緩やかに減少していく)。ただし、海馬破壊群では、error が多い。面白いことに、海馬破壊群の方が、correct reaction time は短くなる。つまり、control 群は正確で遅く、破壊群は不正確で早い。この傾向は、SOC で最も強くなる、という結果。
投稿者 sfujisawa : 17:24 | コメント (23) | トラックバック
2006年04月13日
JNS 4/5(3)
A Biophysically Based Neural Model of Matching Law Behavior: Melioration by Stochastic Synapses
Alireza Soltani and Xiao-Jing Wang
昨日のつづき。
Newsome 論文では、choice probability は、Local な income を反映する、と主張しています(Newsome 論文 Fig 2)。
ここで、「grobal income」は、過去の income がすべて加算されたもの、「local income」は、Fig 2B で示されるような、時定数τをもつ leaky integrator によって過去の income が加算されたもの、を意味します。
ただ、grobal というのは τ→∞ ということなので、τが小さければ local を反映するし、τが大きければ grobal を反映する、ということになります。
それで、この時定数τを求めるのは、choice-triggered-average of rewards (CTA)を用いて調べることができます。(※)
それで、Wang モデルでは、Fig 12 で、 CTA を計算してτを求めています。
ここで、このモデルでのτは、たとえば q+ = q- = 0.06、λtot =0.1(overall biating probabiliy)のとき、τ~15 trial 程度です。
つまり、この Wang モデルでは、すごい local な income が反映されてます。
というのは、この Wang 論文のモデルでは、choice に対する報酬 / 失敗 1 回毎に、LTP/LTD が生じることを仮定しているので、過去の履歴はどんどん上書きされていってしまいます。そのために、local income を強く反映するモデルになっているようです。
ただ、τを決定するのは、1 回の LTP/LTD でのシナプス強度の変化率 q+、qq- なので、もちろんこのモデルでも q+、q- が小さければ、τはどんどん大きくなるはずです。
また、このモデルでは、A or B に selective なニューロン群はそれぞれ 1 つの population しか用意されていませんが、もし、q+ の大きさの違う2つの population があれば、それぞれ local income を表現する群と、 grobal income を表現する群の 2 つの population ができることになります。(あるいは q+ を連続的に分布させることも可能かもしれません。)
ただ、Wang 論文は Newsome 論文に即して書いているので、grobal と local については、そこまで深く議論はしていないようです。
---
(※)
Newsome 2005 J Exp Anal Behav 論文では、サルのデータから計算した CTA は、短い時定数τs と長い時定数τl で fitting できる、と主張しているそうです。
CTA(t) = ωNs exp(-t/τs) + (1 - ω)Nl exp(-t/τl)
これは、pooneil さんが指摘しているような、grobal と local の定量的な分離に相当しているのかも知れません。もっとも、その論文がダウンロードできないのですが。。
投稿者 sfujisawa : 17:03 | コメント (3) | トラックバック
2006年04月12日
JNS 4/5(2)
A Biophysically Based Neural Model of Matching Law Behavior: Melioration by Stochastic Synapses
Alireza Soltani and Xiao-Jing Wang
昨日のつづき。
このモデルでの主張の一つは、reward-dependent の synaptic plasticity を仮定することで、シナプス強度率 cA はリターン RA を表現することが可能である、という点です。
なぜかを簡単に説明します。
まず、ある chioce A に対して 報酬が与えられてたとき、sensory → A のシナプスに LTP が生じて、そのそのときシナプス強度率は
cA(n+1) = cA(n) + q+ [1 - cA(n)]
(記号は昨日のエントリーを参考。)
逆に、ある choice に対して 報酬が与えられなかったときは、 LTD が生じて、
cA(n+1) = cA(n) - q- [1 - cA(n)]
と、このように reward-dependent の synaptic plasticity を生じる、とするのがこの論文でのメインな仮定なワケです。
さてこのとき、もし target i に対する choice probability を Pi とするとき、 ci の変化率は、
Δci = q+ (1 - ci) Ii - q- ci(Pi - Ii)
steady state では Δci = 0 なので、右辺 = 0 としてごちゃごちゃと式変形すると、steady state でのシナプス強度率 ciss が求まります;
ciss = q+ Ri / {(q+ - q-)Ri + q-} 式(7)
これで、もし q+ = q- (choice 成功によって生じる potentiation 率 と 失敗によって生じる depression 率が同じ)なら、ciss = Ri となりますし、あるいは q+ = q- でない場合でも、そこまで大きく違わなければ ciss ~ (q+/q-)Ri と、Ri にたいしてリニアな関係になります。
以上が、シナプス強度率 cA はリターン RA を表現する、という主張です。
つまり、単に reward-dependent な plasticity を仮定するだけで シナプス強度率がリターンを表現するようになるわけで、つまり、biological に Return を表現するのは演算的にはそこまで難しくない、ということを示しています。
---
次に、undermatching phenomena について。
Newsome 論文の Fig 1C を見てみると分かりますが、実際のサルの choice probability は、matching の理論値よりも若干低くなっています。この現象を "undermatching" phenomena と呼ぶらしいのですが、Wang 論文のモデル系でも、この undermatching phenomena が再現されています(Fig 7)。
では、(Wang のモデル系において、)なぜこのような undermatching が生じるのか、Fig 5 で説明しています。
Fig 5 では、横軸は A に対する choice probability、縦軸は、リターン RA(赤)、RB(緑)を示しています。RA と RB が交差する点が、理論的に最適な "matching" 解です。
なぜこのような曲線になるかは、pooneil さんのサイトに mmrl さんによる詳しい解説があります。(全く同じ図が載っています。)
さて、Fig 5B では、RA、RB をそれぞれ cA、cB と置き換えています(上で述べたように q+ = q- なら Ri = ci なので)。この cA と cB を用いて、式(6)のシグモイド曲線より計算した choice probability PA (昨日のエントリ参考)が、Fig 5B の黒線で示されています。横軸の choice probability と、計算された黒線の choice probability PA が一致する点(つまり黒線と点線の交点)が、choice probability になります。
RA (cA) と RB (cB) が交差する matching 点では、cA - cB = 0 なので、黒線の PA は常に 0.5 になります(Fig 3 参照)。(というのは、黒線の PA は cA と cB の差を検出しているので。)なので、黒線と点線の交点、必ず RA と RB の交点よりも低くなります。
これが彼らの説明する、undermatching のメカニズムです。
もう少しおおざっぱに言うと、Biological なシステム(脳)では、matching を行うにあたり、(この論文のように cA と cB を用いているかどうかは分かりませんが、)おそらくリターンの「差」を検出して、そこから演算していると思われます。これが、Biological なシステム では、RA と RB が交差する点(理論的な最適点)よりも低くなる(undermatching が生じる)理由である、ということを言いたいのだと思います。
(つづく。明日は grabal と instantaneous について考えてみたいと思います。)
投稿者 sfujisawa : 19:56 | コメント (0) | トラックバック
2006年04月11日
JNS 4/5
A Biophysically Based Neural Model of Matching Law Behavior: Melioration by Stochastic Synapses
Alireza Soltani and Xiao-Jing Wang
Wang XJ ラボ。
Sugrue、Corrado、Newsome 2004 Science 論文をモデル化した論文。
この Newsome 論文は、 pooneil さんの、言わずと知れたこのシリーズに詳しいです。
Newsome 論文は、pooneil さんのまとめを引用させてもらいますと、「ある選択肢のどちらを選ぶかを決めるときに、人間や他の動物はその選択肢の持つ価値(expected value)によって選択する確率を変化させる。大脳皮質頭頂連合野のLIPニューロンはそのようなexpected valueの比をローカルな時間スケールで計算したものを表象している」ことを主張する論文です。
もう少しややこしく言うと、
LIP ニューロンの(delay-period での)発火率は、Localな income 率(Iklocal/ΣIklocal)を表現している(Newsome 論文 Fig4,5)。Newsome の主張する Local matching law では、Localな income 率は、instantaneous choice probability に等しい(Newsome 論文 Fig 2B)ので、つまり、LFP ニューロンは (instantaneous な) choice probability を表現する
というものです。
それで、今回の Wang 論文の主題は、選択行動における、return Ri、income Ii、choice probability Pi などの行動変数が、LIP 野の local curcuit でどのように表現されうるか、ということことです。この論文での彼らの主張は、LIP ニューロンが Choice probability PA を表現するような local curcuit を構築したとき、
① Choice probability PA は、シナプス強度率の差 cA-cB の関数として表現される。
② Return RA は、シナプス強度率 cA により表現される。
というものです。
---
モデルの構造は以下の図の通り。(Suppl Fig1B より。)
LIP 野を仮定しています。2000 個の integrate-and-fire neuron によるモデル。
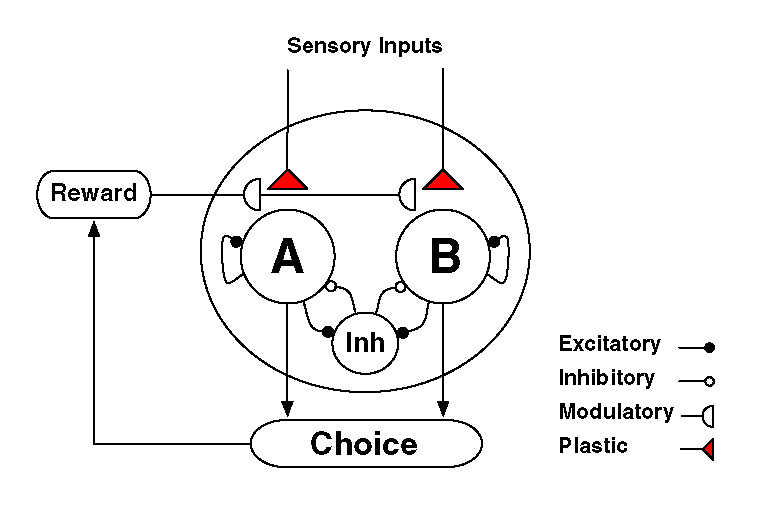
A、B と書かれているのは、それぞれ A、B (たとえば red target、green target)に selective に反応するニューロン群を示します。
仮定は、[sensory input] → [A (or B) に selective に反応するニューロン群] のシナプスに、reward-dependent に、synaptic plasticity が生じる、というものです。
このモデルによって、
(1) LIP neuron の Winner-take-all activity
(2) "undermatching" phenomana
(3) local matching law
などの現象が、シミュレーションにより再現できるようです。
この論文での記号のまとめ。
・行動変数
PA : A を chioce する確率
IA : A を選ぶことにより得られてた income (IA =(A からの Reward)/(total trail))
RA : A からのリターン(RA = IA/PA)
・生理学的変数
g+ : "UP" synapse のコンダクタンス。(g+ = 5.5nS)
g- : "DOWN" synapse のコンダクタンス。(g- = 0.5nS)
q+ : LTP phase において、DOWN synapse が UP synapse に変化する確率
q- : LTD phase において、UP synapse が DOWN synapse に変化する確率
cA : A 群のニューロンに投射するシナプスのうち、"UP"である割合。
cB : B 群のニューロンに投射するシナプスのうち、"UP"である割合。
各シナプスには 2 つの状態があり、"UP"と"DOWN"があるとします。報酬により LTP が生じるとき、q+ の確率で、DOWN シナプスは UP シナプスへと変化するとします。(LTDでは逆。)ここで、cA は、A に投射するシナプスのシナプス強度の率で、cA = 1 なら、A に投射するシナプスがすべて g+ のコンダクタンスを持ち、cA = 0 なら、すべてが g- のコンダクタンスをもつ、ということです。
---
さて、このネットワークモデルは、A、B それぞれの Recurrent による自己増幅系と、A と B の両方の入力を受けて活動する抑制系により、Winner-take-all の persistent activity が実現されます(Figure 1)。(このへんは Wang ラボお得意の話ですね。)
では、cA と cB がどのような関係にあるときに、A 群のニューロンが勝つか?
cA と cB の絶対値には関係なく、cA と cB の差によって決定される、というのがシミュレーション結果です(Figure 3)。これは、以下のような関数(シグモイド関数)で fit されるそうです;
PA = 1/{1+exp(-(cA-cB)/σ)} 式(6)
ここで PA は、A群のニューロンが"Winner"となる確率。この論文では。「A群のニューロンが"Winner"となれば choice A という行動が生じる」という仮定をしているので、PA は「A を chioce する確率」と同義で使っています。
シナプス強度率 cA、cA は、それぞれ choice A、B に対する Return を表現しています(これについては後述)。つまり、PA は、Return の差の関数として表現されることになります。
以上が、上で述べた主張①に相当します。
(つづく)
投稿者 sfujisawa : 09:48 | コメント (10) | トラックバック
2006年04月05日
Neuron 4/5
Temporal Encoding of Place Sequences by Hippocampal Cell Assemblies
George Dragoi and Gyorgy Buzsaki
ブザキラボ。ファーストオーサーのジョージは、3年ほど前に MIT の利根川ラボに移ったらしいので、僕とは在籍期間は全くかぶってない。
それにしても、ややこしくてムズいなぁ、この論文。。
かんたんにいうと、cell assmbly では、 theta 波の 1 cycle という短い時間での Sequence 表現がより精細に可能である、という話。
---
まず、theta 波の 1 cycle の中で表現される Sequence code について説明。
たとえば、論文の図1Aでは、赤色の cell とピンク色の cell では発火率のピークのある場所が約 30 cm 離れている。つまり、ラットが正の方向に進行するとすると、まず赤が発火してからピンクが発火することになるから、cross-correlation を計算すると、そのピークは 0ms からシフトするはずである。(もしラットが 60 cm/sec で走っていたとすると、約 500 msのところにピークが生じるはずである。)
一方、place cell は phase code も行っている。ラットがある place cell の place field に入っていったとき、その place cell の theta 波の中での発火位相は徐々に早い方向にずれていき(phase precession)、発火率がピークとなる場所で theta 中の位相は 180°となる。つまり、たとえば、赤色の cell が発火率ピークとなる場所では、赤色は 180°で発火し、そのときピンク色の cell はそれよりも遅い位相で発火しているはずである。ここで、赤とピンクの cross-correlation を計算すると、1theta cycle(~100ms)以内の場所に、ピークが生じるはずである。
すなわち、二つの place cell(赤とピンク)の cross-correlation を計算すると、大きいピークシフト(数百 ms 単位)をもつ山と、小さいピークシフト(1theta cycle 以内)をもつ山の、二種類の山ができるだろうことが想像できる。
それで、赤 cell とピンク cell の cross-correlation (CCG) を実際に計算したのが Fig 1D である。大きなピークシフトをもつ山(黒い線)に小さな山がのっているのがわかる。
前者を "real-time scale"、後者を "theta-time scale"でのピークシフト、と呼ぶことにする。どちらの場合でも、二つの細胞の place field の距離と、ピークシフトの大きさは、相関することが想像できる。(実際、相関している。前者は Fig 1E、後者は Fig 1G)。
ここで、後者を "theta-time scale"のほうに注目する。こちらの方は、すごい短いタイムスパンのなかで place field の sequence 性が表現されているわけで、興味深いわけです。そこで、このような、theta 波の 1 cycle の中で表現される place cell たちの発火の sequence 性を、"Sequence compression" と名付けることにする。(そしてその place field 距離とCCGシフトの相関の大きさを "Sequence compression index" と名付けることにする。)
(長々と書きましたが、以上の話はもちろんいままで知られてた話です。)
さて、このような 1 theta cycle での Sequence 表現(Sequence compression)のメカニズムは何か?① Theta pacemaker入力によって個々のニューロンが独自に phase precession を起こしているからか?(Pacemaker model)。(たとえば赤 cell とピンク cell が、お互いに全く interaction が無かったとしても、各々の細胞で phase precession さえ生じていれば、Sequence compression は必然的に生じると考えられる)。②それともplace cell 同士が cell assembly として密接に synaptic connection でつながっているから Sequence compression が生じているのか?(Assembly model)。(←Introduction 参照。)
で、どちらが本当のメカニズムなのかは今のところ分かってない。
ここまでが問題提起。
それで、そういうこと(Sequence compression のメカニズム)を調べたい、というのがこの論文の主旨なのだが、それでは何をやったか。
まず、記録した place cell の cell pair について、2つに分類する。
1trail(1lap)(つまり数十秒単位という長い時間スパン)における発火数を観測して、その発火数の変動の correlation が有意に高かった pair を 、"dependent" な place cell の pair、そうでなかった pair を "independent" な pair と分類。つまり、trail-to-trail variability がすごい似た傾向をもつ cell pair を dependentcell pair としている。
このような、"dependent" な place cell の pair では、"independent" な place cell の pair に比較して Sequence compression index が高い(Fig 3C)、というのがメインの結論。
要するに、(このような "dependent" cell pair は、cell assemble を反映しているであろう、と仮定した場合、) cell assmbly では、 1 theta cycle という短い時間での Sequence 表現がより精細に可能である、という話。(つまり、短いタイムスパンでの Temporal coding の信頼性という点では、"Pacemaker model" よりも "Assembly model" のほうが支持できる、という主張。)
問題は、この論文での cell pair の分類("dependent"と"independent"の分類)が、真の cell assembly (あるいは synaptic connenction)をどこまで反映しているか、ということでしょうね。
投稿者 sfujisawa : 19:06 | コメント (3) | トラックバック
2006年03月29日
Nature 3/29
Intellectual ability and cortical development in children and adolescents
P. Shaw, D. Greenstein, J. Lerch, L. Clasen, R. Lenroot, N. Gogtay, A. Evans, J. Rapoportand J. Giedd
これ、なんか衝撃的。少年~青年(7~19歳)の被験者の皮質の厚さを MRI で測定し、IQ との関係を調べた論文。皮質の厚さは成長に伴って変化するらしいが、IQ の高いグループではその変化スピードが速いらしい。また、IQ の高いグループでは、~12歳までは皮質の厚さが増大し、それ以降減少する、という特徴をもつ。
なんか、皮質の厚さの変化スピード、と言われると、すごい genetic に決まってしまいそうな気がします。ただ、この論文では genetic な話は全く触れられていないようです。N&V では、皮質の厚さにおける、genetic な影響を調べた論文と、環境による影響を調べた論文の両方について言及してました。
Reverse replay of behavioural sequences in hippocampal place cells during the awake state
David J. Fosterand Matthew A. Wilson
以前のエントリーで書いたやつ。Moser による N&V あり。
投稿者 sfujisawa : 23:25 | コメント (2)
2006年03月23日
JNS 3/22
Learning-Related Development of Context-Specific Neuronal Responses to Places and Events: The Hippocampal Role in Context Processing
David M. Smith and Sheri J. Y. Mizumori
ラット、place cell。Context dependent に place cell の発火パターンが変化する、という論文。
Context というと、環境の背景色などを指すことが多い。しかし、この論文では、task demand (その task で何が要求されているか)を "context" としている。
ただ、その意味では、Eichenbaum の Nature 1999 や Shapiro の neuron 2003 なども、Context dependent な place cell の発火の論文である。例えば、Shapiro 論文では、context が変化したとき、place field 自体はあまり変化しないが、firing rate が変化する、という主張をしている。
だから、context dependent な plece cell の発火パターン変化だけだと、New ではない。
で、この論文での Task は、十字迷路で、trial によって reward のアームが異なる、というもの(Shapiro のと似ている)。それで、[session 1] では、reward のアームはランダムで、[session 2] では、何 trail かごとに reward のアームが規則的に変化する、というもの。Session 1 と Session 2 を続けて行う。
それで、Session 2 のときのみ、context dependent な place cell の発火が観察された。つまり、学習によってcontext dependent に place cell が発火するようになった、というのがこの論文での主張。
あと、図を見て気になったのは、context dependent に、place field 自体が変化している。
ただ、図をみたかんじでは、なんか place field がきれいでないなぁ。。使ってる電極も、4本対の電極(tetorode)ではなく、2本対の電極だし。。(電極数が少ないと、unit 分離の解像度がどうしても低くなる。)
投稿者 sfujisawa : 19:47 | コメント (0)
2006年03月16日
Neuron 3/2
Tracking Route Progression in the Posterior Parietal Cortex
Douglas A. Nitz
Nitz という人は今まで知らなかった。McNaughton との論文が多いので、McNaughton の弟子かな?
ラット、Parietal Cortex と Hippocampus でのユニット記録。海馬の細胞は、空間における自分に位置に sensitive だが、parietal の細胞は、ある道筋における自分の位置に sensitive である、という内容。
Task は、碁盤目状の課題箱での迷路課題(Fig1)。ところどころ通行止めがあって、通れる pathway は決まっている。ここで、たとえば pathway が、
スタート → 直進 → 右折 → 直進 → 左折 → 直進 → ゴール
というような場合、その各「直進」や「右折」などを segment と呼ぶことにする。
それで、Hippocampus のニューロンは、普通に place field が存在する。一方、Parietal cortex のニューロンでは、place field は存在するが、以下のような特徴をもつ;
・ Pathway dependent である
・ Segment に sensitive である
・ place field の伸縮が可能である
たとえば Fig4A では、はじめの segment に sensitive な parietal neuron の place field が載っているが、「行き」でも「帰り」でも、はじめの segment で発火している。また、pathway が変更されて、はじめの segment がすごく長くなった場合は、それにあわせて place field も大きくなっているのが見てとれる。
---
そうだ、京都にたとえると分かりやすいかも。たとえば、以下の経路を歩くとする;
烏丸七条(京都駅)→ 烏丸五条 → 河原町五条 → 河原町三条
つまり、Segment でいうと、
烏丸通り → 五条通り → 河原町通り
つまり、はじめの Segment に sensitive に反応するニューロンは、「行き」だと烏丸通り、「帰り」だと河原町通りで強く発火する。しかも、伸縮性があるので、もし「行き」のとき五条通りが通行止めで四条通りまで歩かなければいけなくなったとしても、ずっと烏丸通りで発火することになる。
(逆に分かりにくいか?)
---
では、この Parietal と Hippocampus の大きな違いは何か。
まず、Parietal の place field は pathway dependent なので、つまり context dependent だということができる。しかし、context dependent 性だけなら、Hippocampus の place cell にもある。たとえば、Ferbinteanu & Shapiro 2003 では、右に曲がるか左に曲がるかの context の違いにより、place field での発火率が異なる。いってみれば、これも Context dependent である。
しかし、Hippocampus の place cell は、Context dependent ではあっても、あくまで、場所(絶対位置)に sensitive である。一方、parietal では、場所(絶対位置)には independent である(「行き」と「帰り」とでは発火する絶対位置は異なる)。しかも、Parietal では、Segement の大きさにあわせて place field が伸縮する、という実験結果がある。つまり、Parietal の neuron は、ある道筋(どういう手順で目標地点にたどり着くか)における自分の位置に sensitive である、ということである。
投稿者 sfujisawa : 21:22 | コメント (0)
2006年03月07日
PNAS AOP
Relative spike timing in pairs of hippocampal neurons distinguishes the beginning and end of journeys
Matthew L. Shapiro and Janina Ferbinteanu
Shapiro ラボ。PNAS、Edited by Larry Squire。ラット、place cell。
今日のラボミーティングの文献紹介のネタでした。
十字迷路で、スタートアームは異なるけれどゴールアームは同じ、という課題を行います。たとえば、North → West あるいは South → West、という感じで。
それで、ゴールアームで、ほぼ同じ場所をコードしている2つの place cell(cellA、B とする)の発火タイミングの cross correlation をとったところ、North → West あるいは South → West というゴールに至る経路の違いによって、corros-correlation にシフトが生じる、という結果です(Fig 1C)。 (これを journey dependent な cross-correlation のシフトと言っています)。
では、このデータをどう解釈するか。
もし、完全に cell A と cell B のコードする place field が一致していたら、同期して発火することになるので cell A と B の発火タイミングの cross-correlation のピークは t = 0 のところに出てくるはずです。もし、cell A と cell B のコードする place field が少しずれていて、ラットが place field A → place field B の順に走ってきたとすると、当然 cell A のほうが B より早く発火し始めるので、cross-correlation のピークは、t = 0 の点よりずれるはずです。
つまり、place field が cell A と B で相対的にずれたら、その cross-correlation にはシフトが生じるわけです。
ここで、彼らの過去論文を思い出します。Ferbinteanu & Shapiro 2003 Neuron では、同じ十字迷路課題で、West 部に place field をもつ place cell でも、North → West か South → West で、journey dependent に place field が変わりうることを示しています。
つまり、cross-correlation が journey dependent にシフトしているということは、place field が journey dependent に cell A と cell B で相対的にシフトしていることを示唆しているわけです。なので、この論文ではそのjourney dependent な place field の相対シフトが生じているかどうかをきちんと解析しないといけないのですけど、この論文では解析していません。。( journey dependent に place field がシフトしているということを示すと、「それでは内容が Neuron 2003 と同じではないか」、とつっこまれるので、解析していないのでしょうか。)
なんか説明がごちゃごちゃしてしまいましたね。すいません。。
投稿者 sfujisawa : 20:59 | コメント (0)
2006年02月28日
Nature AOP
Reverse replay of behavioural sequences in hippocampal place cells during the awake state
David J. Foster and Matthew A. Wilson
マチュー・ウィルソン。ラット、Place cell ネタです。
今日のラボミーティングの文献紹介で取り上げられたので、ここでも取り上げてみます。
この論文では、Place cell の 発火 sequence が、 ripple 中で逆順になって圧縮される、という驚きのデータを報告しています。
もう少し丁寧にいうと、place cell a が 場所(領域)A をコードしていて、place cell b が場所 B、・・・をコードしているとします。このとき、ラットが場所 A → B → C → ・・・と走ると、Place cell は a → b → c → ・・・という順番で発火することになります。それで、この論文では、ラットが休止しているときに生じている ripple 波の中で、place cell が、・・・→ c → b → a のように逆順に発火する、というものです。しかも、ripple は時間的には 100ms 程度の短いものなので、Sequence は時間的に圧縮されていることになります。
ウィルソンが有名になったのは、おそらくこの論文(Wilson & McNaughton 1994)からだと思うのですが、この以来、ウィルソンは睡眠中での place cell の発火の再現とその Sequence 性にこだわった研究を続けています(Louie & Wilson 2001、Lee & Wilson 2002 など。)ただ、これらの論文では、Sequence の再現は、順方向であると主張しています。Lee & Wilson の Figure 6 に Slow-wave sleep 中の ripple での Sequence 発火が載ってますが、これは順方向です。睡眠中と休憩中での ripple 発生メカニズムがそこまで大きく違うとは思えないのですが、なぜ順方向と逆方向の違いが出てくるのかは今後明らかにしていかないといけないところでしょう。
---
Sharp Wave / Ripple のおさらい。
Sharp wave とは、~100 ms 程度の幅をもつ鋭い波のことで、ripple は 200 hz 程度というとても高い周波数を持つ波です。Sharp wave と ripple は同時に見られることが多いので、Sharp wave / Ripple とよくまとめられます。
実際の波形はこんな感じです。画面真ん中ぐらいにぽこっと上向きにとんがっているのが Sharp wave で、それに乗っている高周波成分が ripple です。(画面は全体で 800 ms)。
Sharp wave/Ripple は、ラットが動いていないとき、および Non REM 睡眠中に観察されます。つまり theta 波が発生していないときです。(ただしこの論文に注意。)Ripple 中では発火数が上昇します。また、ripple 発生源は海馬 CA3-CA1 ネットワークらしいです。
以上の Ripple の特徴より、Behavior 中での place cell の発火活動が、休憩時(海馬への入力が少なくなっているとき)に海馬内で形成される ripple 波により圧縮再現され、これが plastisity を生じさせ、memory consolidation を形成するではないか、という仮説が考えられてきました。
この Ripple 圧縮仮説は、もともとはギューリーがむかし提案したものみたいです(Buzsaki, Brain Res 1986, Neuroscience 1989)。
それで、今回の Wilson 論文では ripple 波により圧縮再現が実際に生じている(ただし逆順で)ことを実験で示したわけですね。
ところで、この Wilson 論文でも、Buzsaki 1986、1989 を引用はしているのですが、彼らは、単なる ripple の説明のところで引用していて、ギューリは「引き方が違うョ!」と言って憮然としてました。。
投稿者 sfujisawa : 23:14 | コメント (0)
2006年02月27日
Nature Neurosci 3月号
The tempotron: a neuron that learns spike timing-based decisions
Robert Gutig & Haim Sompolinsky
Sompolinsky は、以前は Hopfield model などをやっていた理論家です。今回は「Tempotran」という新たな Machine learning を開発した、という話です。
Temporoton では、N 個のニューロンの発火パターンを識別するみたいです。つまり、パーセプトロンに時間情報が入ったみたいなものかな。
識別は、出力ニューロンが時間内に1回以上発火するかどうかで判定します。Fig 1a でいうならば、黒のラスタープロットの入力では出力ニューロンは発火を生じ、灰色のラスターでは発火を生じません。
--
Temporoton では、ある1つのニューロン(出力ニューロン)が、N 個のシナプスの入力を受けているとします。出力ニューロンは、Integrate-and-Fire ニューロンで、
V(t) = Σi ωi Σti K(t - ti) + Vrest (論文中式1)
ωi は i 番目のシナプスの結合強度、K(t) は Postsynaptic potential を表すカーネル関数(Fig1C参照)。
それで、時間 0 から T まで、N 個のシナプスから spike train を入力したとき、出力ニューロンが発火するかどうかでその入力パターンを識別します。
では、学習はどうするか。(Fig 2)。
ある覚えさせたい入力パターンを入力したときに、出力ニューロンの膜電位 V(t) が高かった時間点を tmax を調べます。もし V(tmax) が閾値 Vthr を越えていたら、そのままにします。もし、越えていなかったら、越えるようになるように、シナプス強度 ωi を変更します。
Δωi = λ Σ K(tmax - ti) (論文中式2)
つまり、tmax から時間的に近くに入力があったシナプスのシナプス強度が強くなるよう学習させます。
記憶できるパターンの数は、シナプス数 N の2倍ぐらいだそうです。
---
論文では出力ニューロンの数は1個でしたが、もし出力を M 個にすると、N 入力 M 出力の simple perceptron に似てきます。
では、この Tempotronがパーセプトロンに比べてなにが新しいかというと、
・時間情報を含めたパターンの分離ができる
・演算に Integrate-and-Fire neuron を使っているので、より生理状態に近い。
というところでしょうか。
ただ、問題点としては、
・学習方法ががなんとなく不自然な感じをうける。
・パーセプトロンにくらべて一般性に乏しい。
などが挙げられると思います。
この Tempotron が今後はやるかどうかは、微妙なところかな。。
投稿者 sfujisawa : 21:09 | コメント (0)
2006年02月22日
Nature 2/22
Efficient auditory coding
Evan C. Smithand Michael S. Lewicki
声音などの音の波形は、けっこう複雑な形をしている。
でももし、音の波形が、何個かの関数の足し合わせで表現できたとすると、音を情報として伝達するのに都合がよい。
つまり、音の波形 x(t) を、ある kernel function φ1,・・・, φmを使って表したい、とする。すなわち、
x(t) = Σm Σi smi φm (t - τmi) + ε(t)
と書きたい(論文中式1)。(ε(t) は残差。)では、どのような kernel function を選べば、x(t) を最適に表現できるだろうか?
この問題に対して、音 x(t) が自然界の音であったり、人の声であったりするとき、学習アルゴリズムによって最適な kernel 関数の組をもとめる方法を開発したのが Lewicki NN 2002 論文みたい。これを彼らは「population spike code」と呼んだりしています。
それで、今回の論文では、この最適化学習アルゴリズムによって得られた kernel function と、ネコの auditory nerve の impulse response function(つまり生理学的データ)を比較してみたら、実にそっくりだった(Fig2a、赤(kernal)と青(実データ))、という話。つまり、auditory nerve の impulse response function は、自然界の音や音声などの音情報を伝達するのに、最適な kernel である、という結論。
(この論文では、auditory nerve の impulse response function を「revcor filter」と呼んでいるのですが、このあたりの話は僕は不勉強でよく知りません。。)
補足
Probabilistic Models of the Brain: Perception and Neural Function (eds Rao, Olshausen & Lewicki, MIT Press, 2002) の第12章にも、「population spike code」の話がありますね。
投稿者 sfujisawa : 22:13 | コメント (0)
2006年02月21日
Nature 2/2
GABA regulates synaptic integration of newly generated neurons in the adult brain
Shaoyu Ge, Eyleen L. K. Goh, Kurt A. Sailor, Yasuji Kitabatake, Guo-li Mingand Hongjun Song
今日のラボミーティングでシモルが文献紹介で使った論文。3 週間ぐらい前の論文なのだが、読み落としてたので軽くメモ。
アダルト Dentate Gyrus の新生 granule cell の maturation は、GABA カレントによってモジュレートされている、というはなし。マウス、スライスで、retrovirus を使って DG の proliferating cell に GFP を発現させる手法。
Perforant pathway を刺激すると、まだシナプスを形成していない 3 dpi (days post viral injection) の時点でも、脱分極方向の GABA カレントが流れる(Fig1)。で、GABA なのに脱分極方向に電流がながれるのは、immature の時点では GABA の 反転電位 EGABA が静止膜電位より浅いから(Fig2a)だが、Na-K-2Cl transporter NKCC1 を shRNA で knock down(この辺の手法の話はよく知りません)させると、immature でも EGABA は静止膜電位より浅くはならなかった(Fig2c)。
さらに、この NKCC1 knock down 状態では(つまり分極方向の GABA カレントが流れない状態では)、シナプス形成や dendrite 成長も阻害された、という話。
ちょっとよく分からなかったのは、28 dpi では、GABA の 反転電位 EGABA が既に静止膜電位より深くなっているはず(Fig2a)なのに、なんで Figure 1b では脱分極性の GABA カレントが流れているのだろう?
投稿者 sfujisawa : 20:47 | コメント (0)
2006年02月08日
Nature 2/8
今日、ラボでとなりの席のアソハンがぼくのブログを偶然見つけて喜んでた。何かをググってたらひっかかったみたい。もちろん日本語だから彼は読めないのだけど、それでもちょっと恥ずかしかったり。
ところで、LINUX の firefox って、英語版でもちゃんと日本語文字が表示されるのでちょっとすごい。
Gamma-band synchronization in visual cortex predicts speed of change detection
Thilo Womelsdorf, Pascal Fries, Partha P. Mitraand Robert Desimone
Fries ラボ、Desimone ラボ。サル、V4 での local feild potential とマルチユニット(MUA)測定。データ自体は、Fries et al Science 2001 のときの実験データを使ってるらしく、解析しなおし論文みたい。
Fries 2001 では、attention により V4 での gamma 波が大きくなることを示していたが、今回の論文では、サルの reaction time と gamma 波の大きさ(および gamma-MUA coherence)が高い相関を持っていることを示している。
ところで、Fries の CV を見てみたら、 彼って 29 歳で PI として独立してますね。しかも、MD と PhD 両方持ってるし。どういうこと?
投稿者 sfujisawa : 19:55 | コメント (0)
2006年02月07日
Phys Rev Lett 2/10
Relation between Single Neuron and Population Spiking Statistics and Effects on Network Activity
Hideyuki Cateau and Alex D. Reyes
Reyes ラボの加藤英之さん。Synfire chain を Fokker-Plank 方程式で記述する、という内容。
Fokker-Plank 方程式は、確率密度の時間変化を記述する偏微分方程式です。
例えば t=0 で x=0 の点にいる粒子のブラウン運動を考えてみます。t=0 だと粒子の位置の確率密度分布はデルタ関数を用いて、p(x) = δ(x) とあらわされるでしょうが、t が大きくなるに従って次第に平べったい紡錘型の分布になることが想像できると思います。この p(x,t) を記述するのが Fokker-Planck 方程式です。(式の形は拡散方程式に似ています。)
それで、Reyes 2003 NN 論文の現象を数理的に記述するのが本論文の内容です。具体的には、N 個のニューロンを含む各 Layer での膜電位 Vm の確率分布 p(Vm, t) をFokker-Planck で記述するのが目的です。
(↑Reyes 論文は、ニューロンが synfire 的に feed-forward でつながっているとき、Layer 1 に Gaussian noise 入力を入れても、後方の Layer では Synfire 効果によりリズミックな出力が観察される、という論文。)
それで、今回の論文のポイントは、ニューロンの発火が、Poisson 過程ではないと仮定したところ。
これは、たぶん、Poisson 過程を仮定すると、うまくいかなかったためだと思う。何でうまくいかないかというと、ニューロンが Poisson 過程で発火するとすると、その inter-spike interval(ISI)は、指数分布に従う。つまり、t=0 のとこにピークが出るような分布。これを使うと、Fokker-Planck 方程式を解いたときに、Reyes 実験のような繰り替えし構造が出なかったのだと思う。(たぶん。)
(モデル細胞に、NMDA channel や NaP channel (persistent sodium channel) のような持続性のカレントを仮定すれば、たぶん解決できるのだろうが、ただ、それでは理論としての一般性がどうしても弱くなるので避けたと思われる。この論文では、モデル細胞にはもっともシンプルな leaky integrate-and-fire neuron を用いている。)
それで、neuron の ISI の分布形状を、指数分布ではなくガンマ分布であると仮定した。ISI 分布がガンマ分布であると仮定すると、その分布形状は実際のニューロンの ISI 分布と似たものになる。(たとえば 論文 Fig1a-middle)。
(ガンマ分布 Γ(α,σ) は α=1 のとき指数分布となることに注意。)
その仮定のもとで得られた Fokker-Planck 方程式が、式9(ノイズ入力が Gaussian noise のとき)あるいは式13(ノイズ入力が Gaussian noise でないとき)というわけです。
要約すると、各ニューロンの発火の ISI を、指数分布(←発火が Poisson 過程の場合)ではなくガンマ分布で近似すると、Reyes 論文の現象をうまく Fokker-Plank 方程式で記述できる、ということです。
投稿者 sfujisawa : 19:15 | コメント (0)
2006年02月06日
Neuron 2/2
Object Selectivity of Local Field Potentials and Spikes in the Macaque Inferior Temporal Cortex
Gabriel Kreiman, Chou P. Hung, Alexander Kraskov, Rodrigo Quian Quiroga, Tomaso Poggio, and James J. DiCarlo
Poggio が著者に入ってる。マカク IT 野での電気生理。Local field potential (LFP)の Object slectivity と、発火活動(おもに multi-unit activity)との関連について。(multi-unit activity とは、スパイクを単細胞に分離してなくてごちゃ混ぜの状態のこと。)LFP はその領域への "input" を反映していて、spike は "output" を反映している、という観点で書かれている。
IT 野は object selective なニューロンが存在する部位だが、LFP にも object selectivity がある程度存在するということを示した論文。
ぼく的には Figure6 がけっこう好き。LFP はどのくらいの距離内の発火活動に影響を与えているか、の算出をしている。電極を 1mm 毎に 10 本並べて(それぞれを電極1,電極2、・・とする)LFP と マルチユニットを記録しているのだが、その LFP と マルチユニットの correlation と距離の関係を解析している。具体的には、例えば、[Cutoff Distance = 3 mm] の場合、
「average of nearby sites (黒点)」では、電極1で記録された LFP と、電極1&2&3で記録されたマルチユニットの平均、との Correlation を計算する。
「avarage of remote sites(白点)」では、電極1の LFP と、電極4~10のマルチユニットの平均、との Correlation を計算する。それで、この2本の線が交わるのがだいたい 3 mm のところ。これより、LFP は 3mm 程度の距離内の細胞の発火(の平均)と関係している、ということを示唆してる。
投稿者 sfujisawa : 23:33 | コメント (0)
2006年02月03日
Science 2/3
最近ちょっと日記をさぼり気味。
WangXJの先週のJNS論文について書きたいのだけど、なんかうまく書けない。まあそのうち。
Rats Smell in Stereo
Raghav Rajan, James P. Clement, and Upinder S. Bhalla
匂い源の位置の特定に関する実験(Rat)。Rat での odor-localization task など。鼻を穴につっこむと右と左から空気が吹き付けられて、そのどちらかは匂い付き。右から空気が匂い付きなら右のノズルから水(報酬)がもらえて、左からの空気が匂い付きなら左から水がもらえるという task 。この task を、ラットは約6日の training で80~90%の正解率でできるみたい。あと、嗅球でのユニット記録もやっている(これは麻酔下)。嗅球では匂いの derection-selective な細胞がいるみたい。つまり、嗅球の時点ですでに匂い源の方向の分離ができているという話。
投稿者 sfujisawa : 23:30 | コメント (0)
2006年01月20日
JNS 1/19
Spike Count Reliability and the Poisson Hypothesis
Asohan Amarasingham, Ting-Li Chen, Stuart Geman, Matthew T. Harrison, and David L. Sheinberg
First author のアソハンは、今はうちのラボのポスドクで、実は席が僕のとなり。数学出身で、専門は確率統計みたいです。僕は統計数理は苦手なので普段はこの手の論文はパスなのですが、pooneil さんのとこでも紹介されてますし、何より席がとなりなのでこの論文はさすがに読まないとまずいですね。。(笑)
ストーリーを要約すると、
(1) ニューロン発火のスパイク列は、統計的に分散が大きいので、Poisson process ではないかと考えられていた。
(2) しかし、今までの統計方法では、分散を大きく見積もりすぎている。
(3) そこで、Poisson 過程であるかどうかを判定する新しい統計手法を開発した。
(4) この統計手法で、実際のサル電気生理から得られたスパイク列のデータを解析すると、Poisson 過程よりも reliable なものであることが分かった
という内容。
では、まず Poisson 分布について。
たとえば、あるニューロンの発火について考えてみます。
ある短い time bin ΔT(例えば 1ms)で、ニューロンが発火する確率 pΔT は非常に小さい(例えば pΔT = 0.002)が、観測時間 n×ΔT が長い(例えば n = 1000)と、期待値 p×n はあるそこそこの値をもつ。(ここでは p×n=0.002×1000=2、つまり 2 Hz)。このニューロンの発火を観測した場合に、観測された発火数 が例えば 3 である確率は、pΔT が一定とするならば二項分布に従うので、
1000C3 0.0023(1-0.002)997。この計算を直接実行するのは大変である。しかし、p が非常に小さく、n が非常に大きい場合、観測される発火数 m の確率を求めるのには以下の定理が使える(ポアソンの小数の法則);
nCm pm(1-p)n-m → e-λλm/m!
ここでλは期待値(λ=np)。これが Poisson 分布で、つまり Poisson 分布は二項分布の n → ∞、p → 0 の極限での分布のこと。(また、λを大きくしていくと、ポアソン分布は正規分布に近づく。)
この式から上の確率を求めると、λ=2,m=3より、e-λλm/m!=0.18。
(以上の話は「統計学入門」(東大出版会)より改変)。
ポイントは、微小 time bin ΔT における発火確率 pΔT が、常に一定であるというのが、Poisson 分布を持つための必要条件である。
ところで、現実のニューロンでは、pΔT は、長い時間のスケールで見ると、一定でないように見える。ある event があれば発火率は上昇するだろうし、何もなければ発火率は減少するかもしれない。つまり、発火率 λ は時間によって変化する(すなわち、λ=λ(t)、pΔT = pΔT(t) )。ただし、λや pΔT が大きなスケール時間的に変動しても、局所的(比較的短い時間スケール)でみれば、 pΔT は、すべての time bin に対して等価である、としたのが、inhomogenous Poisson process 。(このへんの話は、Shadlen & Newsome 1999 のあたりで提案された話らしいのですが、僕はあの有名論文、難解なためにいつも途中で挫折してしまっていまだ理解できておりません。。)
つまり、Poisson Process でない、ということは、局所的に見ても、pΔT は、すべての time bin に対して等価でないということであります。
* * *
ここで、本題に入る前に、以後使用する記号をまとめておきます:
i 番目の trial で観測された発火率 mi
n 回の trial で観測された発火率の平均 μ'= (1/n)Σmi
n 回の trial で観測された発火率の分散 σ'2
真の発火率(発火率の期待値) λ
真の分散 σ2
(真の値は unknown なので、観測によって得られた値で代用する、ということです)
さて、ニューロンの spike train が Poisson process に従う、という根拠は、「trial毎の発火率の分散が大きい」という実験事実があるためです。
Poisson 分布では、その数学的性質上、期待値と分散は等しくなります。つまり、Poisson 分布では、Fano factor = σ2/λ = 1。
Shadlen 論文での主張は、観測によって得られた Fano Factor(σ'2/μ' )が 1~1.5 になるので、spike train が Poisson prcess であるとの仮説が成り立つのではないか、というものです。
それで、ここからが本題になるのですが、以上の Shadlen 論文の主張に異議を述べたのが、本論文です。
このような spike 列の統計的な処理を行うときには、「trial毎の観測の揺らぎは存在しない(trail-to-trial statistical stationarity)」という暗黙の仮定をおいています。
つまり、観測による trial毎の揺らぎを考慮していません。しかし、もし、trial毎に揺らいでいるのであれば、Fanofactor(=分散/平均)が大きい値を持つのは当然、ということになります。
ここで「観測の揺らぎ」と言っているのは、観測自体に内包しているしているような揺らぎのことです。さいころを振るような trial の場合は、tiral毎の観測の揺らぎはないと考えられます。しかし、サルの電気生理のような場合、trail毎の条件が同じであったとしても、サルの集中力や疲れなどの hidden な条件によって揺らぎが生じるので、trail毎の観測の揺らぎが乗ってしまう可能性があります。
つまり、trial毎に揺らぎが考えられる観測では、単に σ'2/μ' が 1~1.5 になるからといってそれが Poisson Process であることの主張とするには根拠が弱い、という主張です。(なぜならσ'にはニューロン発火の純粋な揺らぎ(真の分散)に加えて、サルの集中力といった観測の揺らぎといういらないものが入ってしまうから。)
それで、trail 毎に揺らぎがある場合でも Poisson Process であるかどうかを判定する方法を提案したのが、本論文なわけです。
ではどうやるか。
この論文での戦略として、帰無仮説を立ててそれを棄却できるかどうかで判定するという手法を用います。
帰無仮説(null hypothesis)H0
m1, m2, ・・・, mn
は独立な Poisson random value である
で、この帰無仮説を棄却できれば、Poisson 過程ではないと言える。(棄却できなければ Poisson 過程であるともないとも言えない)。
で、その判定として、以下の不等式を用いる;
Σ mi2 ≦ f (論文中の式(2))
つまり、帰無仮説を棄却できる値 f を見つけることができれば、統計的に Poisson 過程でないかどうかの判定が可能になる、というわけです。それで、この値 f を数学的に見つけてやろう、というのがこの論文の前半での主題。
f は、trail回数 n、観測平均 μ'、および棄却のレベル(どれくらいの確率で棄却できるかということ)αの関数になると考えられる。すなわち、f = f(n,α,μ')。
それで、結局、論文中の式(10)によって f を求めることができる、というのが本論文での結果です。
(式(10)の導出方法の詳細は省略しますが、Trial-to-trial statonarity を仮定した場合との統計的な比較によるみたいです。 )
数値的には、モンテカルロシミュレーションを使って、f を求めるみたいです。
一応、ホームページに f を求める Matlab プログラムを置いているみたいなので、そのプログラムを使えば、自分のデータを試してみることも可能です。
以上の話をまとめると、
(0) 今までは、Poisson process かどうかの判定に、Fano factor(分散/平均)が~1であるかどうかを使っていた。
(1) しかし、Trial-to-trial に揺らぎ(観測に依る揺らぎ)がある場合は、Fano factor が真の値より大きくなってしまうという不都合がある。
(2) それで、新しい統計手法を開発した。ついでに、簡単に使えるようプログラム化した。
ということです。
それで、論文の後半では、実際のサル電気生理での spike 列のデータを使って、Poisson 過程であるか否かを解析しています。結果を簡単にいうと、stimulus onset から400msぐらいまでの区間(発火率が比較的高い区間)では、スパイク列は Poission 過程ではない、(Poisson よりもっと規則正しい)ことが統計的に示されています(Figure4b)。Stim onset から 500 ms以上離れてくると、Poisson 過程でないと言えなくなってきます(帰無仮説が棄却されないため)。
* * *
さて、(上で述べたことを繰り返しますが、)Poisson Process でない、ということは、局所的に見ても、pΔT は、すべての time bin に対して等価でないということです。
端的に言えば 、ニューロンには、refractory effect(1回発火した直後にはすぐ(~2ms)には発火できない特性)や burst などの biophysical な特性があるが、発火率が上昇すれば上昇するほど、そういう特性に支配されて、ランダム性(Poisson 性)が減少していく。まあ、当たり前といえば当たり前の話ですが、統計数理的に、どのレベルで Poisson 的であるかそうでないかを議論したのが、この論文の新規性ということでしょう。
投稿者 sfujisawa : 19:42 | コメント (2)
2006年01月12日
JNS 1/12
Head Direction Cell Representations Maintain Internal Coherence during Conflicting Proximal and Distal Cue Rotations: Comparison with Hippocampal Place Cells
D. Yoganarasimha, Xintian Yu, and James J. Knierim
Knierim ラボ。ラット、head direcion cell(thalamus からのユニット記録)。
Head derection cell の発火は Local cue によるか Distal cue によるか、について調べた論文(このエントリ参考)で、Knierim お得意の、Distal cue を 時計回りに回転させ、 Local cue を反時計回りに回転させ、place field がどっちに回転するかで調べる手法。
で結果は、ほとんど(94%)の dead derection cell の head direction field が、時計回りの方向に回転した。つまり、thalamus の Head direction cell は、distal cue によってその head direction field を形成している、という話。この内容、2年前の SfN で見かけた記憶がある。
The Continuous Wagon Wheel Illusion Is Associated with Changes in Electroencephalogram Power at ~13 Hz
Rufin VanRullen, Leila Reddy, and Christof Koch
Koch ラボ。高速で回転しているタイヤを見ていると、回転している方向とは逆の方向に回転しているように見えることがある。これを「Wagon wheel illusion」と呼ぶらしい。
ビデオの映像(例えば車のコマーシャルとかかな)で Wagon wheel illusion を生じる場合は、ビデオの sampling rate が wheel の回転周波数とほんの少しずれることによって説明できるが、この現象は、ビデオではない現実の視覚でも生じる(例えば独楽を見ているときなどかな)。この wagon wheel illusion は、wheelが 10~15 Hzで回転しているときに生じるらしい。で、このメカニズムは今のところ分かっていないらしい。
今回の論文では、被験者が、wagon wheel illusion を感じているときに脳波を測定したところ、~13 Hz 成分(α波成分)の減少が観察された、というもの。それで、この周波数の脳波が、wheel rotation の知覚に関係している、という話。Illusion を感じるときの wheel の回転周波数と脳波の周波数が一致しているのが、偶然なのか何か意味があるのかは分からない。
Tactile Spatial Attention Enhances Gamma-Band Activity in Somatosensory Cortex and Reduces Low-Frequency Activity in Parieto-Occipital Areas
Markus Bauer, Robert Oostenveld, Maarten Peeters, and Pascal Fries
Fries ラボ。
投稿者 sfujisawa : 21:49 | コメント (2)
2006年01月05日
Neuron 1/4
Place-Selective Firing of CA1 Pyramidal Cells during Sharp Wave/Ripple Network Patterns in Exploratory Behavior
Joseph O'Neill, Timothy Senior, and Jozsef Csicsvari
Csicsvari ラボ("チチバリー"と読むらしい)。海馬で Sharp Wave/Ripple が発生するのは、ラットが静止しているときか、NREM睡眠中。ラット歩行時は、Theta 波が発生する。この論文では、SPW/Ripple が、ラット歩行時にも発生する(つまり、theta 波と同時に出現したりする)という現象を発見した、というもの。しかも、この ripple 中での pyramidal cell の発火も、place-selectivity がある、という内容。まあ、ブザキラボとしては ripple が memory consolidation に関わっているのではないかという漠然とした仮説を持っているわけで、睡眠/静止時だけではなく歩行時にも ripple が発生するという現象は、興味深いわけです。
ところで、Jozsef Csicsvari は、ブザキラボの出身。はじさんとか Ken Harris と同じぐらいの世代('98~'02ぐらいを中心にブザキラボのポスドクだった世代)。この世代の人たちは優秀で、はじさんもケンもジョゼフも、みんなまだ若いのすでに独立して自分のラボもっているのだから、スゴいものである。
Shunting Inhibition Improves Robustness of Gamma Oscillations in Hippocampal Interneuron Networks by Homogenizing Firing Rates
Imre Vida, Marlene Bartos, and Peter Jonas
Threshold Behavior in the Initiation of Hippocampal Population Bursts
Liset Menendez de la Prida, Gilles Huberfeld, Ivan Cohen, and Richard Miles
Enhancement of Spike-Timing Precision by Autaptic Transmission in Neocortical Inhibitory Interneurons
Alberto Bacci and John R. Huguenard
Predictive Neural Coding of Reward Preference Involves Dissociable Responses in Human Ventral Midbrain and Ventral Striatum
John P. O'Doherty, Tony W. Buchanan, Ben Seymour, and Raymond J. Dolan
投稿者 sfujisawa : 21:26 | コメント (0)
2005年12月23日
Nature Rev Neurosci 12月号
Neuroscience nanotechnology: progress, opportunities and challenges
Gabriel A. Silva
ニューロサイエンスとナノテクについての Review。先端技術については勉強しとかないといけないなー、と常々思っているので、とりあえず読んでみました。この Review で紹介されたあったのは、おもに以下の技術について(table1より)。(3)-(4)のあたりが、この著者の専門みたい。
(1) Molecular deposition & Nano-lithographic patterning
リソグラフィー(露光技術。半導体の回路パターンなどを作るときに使う)や有機分子の蒸着技術などを応用して、なにか bio-interactive なパターン回路をナノメーターの解像度で作ってやろうというもの。
(2) Molecularly functionalized surface
基質の表面に、なにか molecular をつないでやって、その molecular の評価系みたいなものをつくろうというはなし。
(3) 量子ドット
量子ドットを、蛍光物質として使う。退色が少なく、鋭い emission spectrum をもつという特長があるらしい。この量子ドットになにか自分の好きな分子をくっつけてやって、イメージングに使う、というもの。いま流行ってるみたい。
(4) 自己組織的に形成される nanotube
疎水性の炭素のしっぽと、親水性のペプチドからなる分子(脂肪みたいな感じかな)を作製する。このペプチド部分は、たとえば、IKVAVという分化を促進するような物質にしてやる。この分子は、ある条件下で自己組織的に集積して、nanotube を形成し、巨視的にはゲル状となる。で、このゲルのなかに projenitor cell を入れて飼ってやると、neuron への分化が促進されるとか。(文献 21)
(5) カーボン・フラーレンをベースとした化合物
Anti-oxidant としての働きをするらしい。
(6) Blood-brain barrier を通過する nanoparticle
drug delivery に使う。
--
量子ドットというのは、最近よく耳にするのですが、僕は何のことか知りませんでした。今日 Wikipedia で調べたら、こんな感じに書いてました:
「主に半導体において、結晶成長や微細加工により電子の持つド・ブロイ波長(数nm~20nm、nmは1x10-9m)の粒状の構造を作ると、電子はその領域に閉じこめられ電子の状態密度は離散化される。閉じ込め方向を1次元にしたものを量子井戸構造、2次元のものを量子細線、そして3次元全ての方向から閉じ込めたものを、量子ドットと呼ぶ。」
ん?3次元すべての方向で電子を閉じこめるって、どういうことじゃろー?このページを読んだ感じでは、つまり結晶のサイズを電子のドブロイ波長程度にすれば、それだけで電子は閉じこめられてしまうみたいね。
簡単に計算してみます。
例えば、半導体材料として一般的な、GaAs (ガリウム砒素) の結晶の場合、1.43 eV 以上のエネルギーを与えると、電子は価電子帯からバンドギャップを飛び越え、伝導バンドへ移動する。で、普通であれば伝導バンドでは、電子のとりうるエネルギーは自由。
ここで、1.43eV での電子のドブロイ波長は、計算してみると、約 1 nm 。もし GaAs の結晶サイズ d が、3次元ともに 1 nm 程度であれば、電子は、伝導バンドでも、波長が d (の整数分の1)に拘束されてしまって、つまりとりうるエネルギーが離散的になる、というわけですね?(間違ってたらすいません。)
つまり、GaAs なり、CdSe なりの半導体の結晶のサイズをナノサイズの大きさにすればそれが量子ドットになるわけで、蛍光物質として使う場合は、結晶が壊れないかぎり退色しないし、発光のスペクトルも鋭くなるわけですね。難しいのは、粒子サイズをそろえること(←粒子サイズによってスペクトルが変わってくるので)と、粒子に自分の好きな有機分子をくっつけることかな?
量子ドットはもともとは、単一電子トランジスタやフォトニクス素子、レーザーなどへの応用に期待がもたれている材料みたいですね。
投稿者 sfujisawa : 23:34 | コメント (0)
2005年12月16日
Science 12/16
今日は午後から、うちの学科で Holiday party なるものがあった。(でも僕は作業が終わらなくて出れなかった。。)ということは、一般的には、明日からがクリスマス休暇なのかな?
Long-Term Modulation of Electrical Synapses in the Mammalian Thalamus
Carole E. Landisman and Barry W. Connors
Electrical synapse でも、mGluR 経由で、長期減弱が起こるらしい。。
系は、ラット、thalamocortical slice 、thalamo-reticular neuron (Interneuron) のパッチクランプ。
投稿者 sfujisawa : 22:48 | コメント (0)
2005年12月12日
Neuron 12/8
メモ。
Epileptogenic Actions of GABA and Fast Oscillations in the Developing Hippocampus
Ilgam Khalilov, Michel Le Van Quyen, Henri Gozlan, and Yehezkel Ben-Ari
ベン・アリ研。ラット、スライス系の電気生理。幼年期のGABAは興奮性に働くことが知られているが、この興奮性のGABA系が、幼年期の epileptogenesis に関わっている、という話。
Internal Dynamics Determine the Cortical Response to Thalamic Stimulation
Jason N. MacLean, Brendon O. Watson, Gloster B. Aaron, and Rafael Yuste
ユステ研。Thalamus 付きの cortex スライスでの、カルシウムイメージ実験。Cortex において、自発的な活動(UP/DOWN)と、thalamusを刺激したときに生じる活動の類似性を議論した論文。
Active Maintenance of Associative Mnemonic Signal in Monkey Inferior Temporal Cortex
Masaki Takeda, Yuji Naya, Ryoko Fujimichi, Daigo Takeuchi, and Yasushi Miyashita
宮下研。これはちょうど、今年のSfNで発表されてたやつですね。ちょうどその場にいらした納家先生に解説していただきました。
Two Cortical Systems for Reaching in Central and Peripheral Vision
Jerome Prado, Simon Clavagnier, Helene Otzenberger, Christian Scheiber, Henry Kennedy, and Marie-Therese Perenin
投稿者 sfujisawa : 22:42 | コメント (2)
2005年12月07日
JNS 12/7
メモ。
Recognition of Familiar Individuals in Golden Hamsters: A New Method and Functional Neuroanatomy
Wen-Sung Lai, Leora-Leigh R. Ramiro, Helena A. Yu, and Robert E. Johnston
ハムスターを用いた social memory の論文。動物(rodent)の個体識別において、相手のことを知っている(familiar)かどうかの記憶だけでなく、相手の性質までも記憶しているかどうかを評価する系を考えた、という論文。
被験者ハムスターは、experience phase において、2匹のハムスター(ハムスターAとハムスターBとする)とそれぞれ面会する。面会の仕方は、ハムスターAとは、金網を隔てて穏やかに面会させる。ハムスターBとは、戦わせる。(ハムスターBは敢えて喧嘩の強い奴を選んでいるので、被験者ハムスターは、ハムスターBには負ける。)
test phase では、被験者ハムスターは、Y-maze を行う。ここで、Y-maze のうちの一本のアームに、ハムスターA、あるいは、ハムスターBがいる。で、被験者ハムスターが、その他人ハムスターのいるアームをどれだけ避けるか、で評価。被験者ハムスターは、ハムスターBがいるばあい、そのアームを有意に避ける、という結果。
あと、c-Fos 実験や、局麻実験など。海馬依存らしい。
Materials&Methodsを読むかぎり、評価系にはあまり洗練されてない部分も多くあるように感じたが、でもまあ面白い論文かな。
ところで、この論文で参照してあった、この論文が面白かった。羊も(羊の)顔認識ができるらしい。しかも、50匹分の羊の顔写真を識別できて、それを2年以上も記憶しているとか。。
Motor Memory Consolidation in Sleep Shapes More Effective Neuronal Representations
Stefan Fischer, Matthias F. Nitschke, Uwe H. Melchert, Christian Erdmann, and Jan Born
Competitive Stimulus Interactions within Single Response Fields of Superior Colliculus Neurons
Xiaobing Li and Michele A. Basso
Neuronal Sensitivity to Microsecond Time Disparities in the Electrosensory System of Gymnarchus niloticus
Atsuko Matsushita and Masashi Kawasaki
投稿者 sfujisawa : 22:03 | コメント (0)
2005年12月01日
Nature Neurosci 12月号 (2)
メモ。つづき。
Neuronal correlates of subjective sensory experience
Victor de Lafuente & Ranulfo Romo
Romo。例によって、サルの手に振動刺激をあたえる実験。今回は、振動刺激の強さを、感じるか感じないかの閾値付近でランダムに変化させる、というもの。サルは、刺激を感じたら「Yes」ボタンを、感じなかったら「No」ボタンを押す、という課題。記録は、Primary somatosensory cortex (S1) と、medial premoter cortex (MPC) から。刺激の閾値付近では、MPC のニューロンの発火はサルの回答とよく相関しているが、S1 ニューロンの発火はサルの回答と相関していない、という結果。で、MPCは、主観的な感覚経験をコードしている、というのが彼らの主張。
Cognitive control mechanisms resolve conflict through cortical amplification of task-relevant information
Tobias Egner & Joy Hirsch
ヒト、fMRI。Attention は、taskに関連する情報を増幅すべく働くのか、taskに関係しない情報を抑制すべく働くのか、あるいはその両方なのか、ということに焦点をあてた論文。課題は、改変型 Stroop task。ある著名人(政治家 or 俳優)の顔写真の上に、別の著名人(政治家 or 俳優)の名前が文字で書かれた画像を見て、俳優か政治家かを選ぶ課題。(たとえば、ロバート・デニーロの顔写真の上に、「毛沢東」と文字が大きく書かれた画像。)顔写真から俳優か政治家を選ぶ課題と、文字から選ぶ課題の両方を行う。
このとき、fusiform face area (FFA) での BOLD 信号を測定。FFA は face processing に関係するといわれている領域らしい。
結果は、FFAでは、顔写真から選ぶ課題の時、顔写真と文字の属性が違う場合(政治家+俳優 or 俳優+政治家)は、属性が同じ場合(政治家+政治家 or 俳優+俳優)よりも、活動が上昇する。
一方、文字から選ぶ課題の時(つまり顔写真は distractor)、顔写真と文字の属性が違う場合は、属性が同じ場合よりも、活動が減少しない。
つまり、Attention は、taskに関連する情報を増幅すべく働く、という結論。
投稿者 sfujisawa : 20:29 | コメント (3)
2005年11月30日
Nature Neurosci 12月号
なんか、Peter Dayan が2報も載ってますね。。
Matching storage and recall: hippocampal spike timing–dependent plasticity and phase response curves
Mate Lengyel, Jeehyun Kwag, Ole Paulsen & Peter Dayan
理論(ちょっと実験付き)。連想記憶の encoding と retrieval について。ある記憶のパターンが、ニューロンの発火 phase により表現されるとする。Encoding過程において、シナプス重み wij が、spike-timing dependent plasticity によってモジュレートされるとする。Retrival過程においては、pre の発火が、post の発火 タイミング に対して影響を与えることによって、記憶されたニューロン群の発火パターンを誘起する。さて、ここで、pre の発火が post の発火 タイミング に対してどのように影響の与えると、encodeしたパターンをもっとも最適に誘起できるか、というのを理論的に導出した、という内容。
* * *
N個のニューロンのネットワークを考える。ネットワークのシナプス重み分布は wij とする。情報は、ニューロンの発火タイミングによってコードされるとする。M個のパターンを記憶するとする。
x1 , x2 , ・・・ , xM
ここで、xim は、m 番目のパターンにおける、i 番目のニューロンの発火タイミングを示す。(太字はベクトル。)( Hopfield Network では、xim は0か1かのバイナリを考えるが、この論文では、ある基準時間からの時間差を考える。)
ある cue 入力 x' によって、パターン xm が誘起されたら、連想記憶が形成されていることになる。
ここで、x', wij に対するパターンxの誘起確率
P[ x | x', wij ] (x ∈ {xm})
を最大化するような retrival はどのように記述されるか、という考え方の枠組み。
で、彼らの結論は、encoding 過程において、シナプス重み wij の変化が、ある関数 Ω によって
Δwijm = Ω(xim, xjm)
と記述される場合、Retrieval 過程において、上述のような P を最適化するように retrive するには、pre の発火があたえるpost の発火 タイミング に対しての影響が以下のような関数で記述されればいいらしい;
H(xi , xj) = wij ・ ∂xi (Ω(xi , xj)) (論文中、式(3))
つまり、Ωの微分に比例。
たとえば、Encoding が Fig.1b のような関数によってなされる場合、retrieve は、その微分をとって、Fig1c のような関数でなされるのが最適、ということらしい。このような関数を、彼らは "phase response curve" と呼んでいる。
で、海馬ニューロンでは、intrinsic property として、このような phase response 特性をもっている、ということを実験的に示してます。
うーん、それにしても Dayan の論文はムズい。。なんかわかったようなわからんような論文ですね。。
投稿者 sfujisawa : 19:29 | コメント (0)
2005年11月03日
Nature Neurosci 11月号 追記
昨日の ripple 論文、ギューリーが N&V を書いてたんですね。(pooneil さんのエントリを見てはじめて気づきました。。)まあ確かに、ギューリーにとってはハマリ論文ですね。。
さっそく読んでみました。このN&Vでは ripple と consolidationとの関連について焦点があてられているみたいですね。。
① ripple が休憩中/睡眠中(NREM)に生じること、② 睡眠前の経験(行動課題)によって ripple が shape されること(※)、③ ripple は海馬で発生して、周辺皮質(ECなど)に伝播すること、などから、(状況証拠的に)ripple は睡眠中の memory consolidation に対してなんらかの役割(たとえば、機能的 neuronal assembly の形成など)があるのではないか、と考えられている。
で、今回の論文では、スライス系で、シナプス可塑性と ripple が絡んでいるよー、
という話の流れのNews&Viewsみたいでした。
もっとも、今回の論文では、シナプス可塑性を生じさせる(ぐらい強い)刺激でrippleが発生した、という報告なので、シナプス可塑性と ripple の生成とはパラレルな話なのですが、まあでも、その関連性を示唆するデータということで面白いのでしょう。
(※) ギューリーの N&V では、ここで Wilson&McNaughton Science 1994 の論文を参照してました。行動課題中に、spike の correlation が高かった Place cell のペア(つまりplace filed が overlap している place cell の pair)は、その課題後の睡眠中でも(課題前の睡眠に比べて)spike の correlation が高い、ということを示した、あの有名な論文。
ん?あの論文に ripple のことなんて書いてあったっけ?、と思って読み返してみたところ、確かに後ろの方に書いてました。「correlation は ripple 中の方が、ripple 外のときよりも有意に高い」らしいです。(でも、課題前の睡眠での ripple と、課題後の睡眠での ripple での違いまではこの論文では調べてないようです。)
投稿者 sfujisawa : 19:36 | コメント (0)
2005年11月02日
Nature Neurosci 11月号
メモ。
Induction of sharp wave–ripple complexes in vitro and reorganization of hippocampal networks
Christoph J Behrens, Leander P van den Boom, Livia de Hoz, Alon Friedman & Uwe Heinemann
海馬スライス系でのRipple波の生成の報告。
Ripple波(100Hz程度)は海馬特有の脳波で、ラットが動いていないときとSlow-wave sleepの時に見られる。ラットの慢性実験をやっているときは、Rippleのありなしで、電極が海馬に到達したがどうかが確認できる。しかも海馬のレイヤーによって波形が違うから、電極がどのレイヤーにあるのかまでがだいたい特定できるために、Rippleはけっこう重宝する。
今回の論文は、海馬スライス系で、シャッファーに高周波刺激を5分おきに何回も繰り返していると、自発的に CA3 で Ripple波 が見えるようになる、という報告。つまり、Rippleの起源は CA1-CA3 networkということですね。
Fine-scale specificity of cortical networks depends on inhibitory cell type and connectivity
Yumiko Yoshimura & Edward M Callaway
キャラウェイ研の吉村さん。この論文の続報かな。
Activity-dependent decrease of excitability in rat hippocampal neurons through increases in Ih
Yuan Fan, Desdemona Fricker, Darrin H Brager, Xixi Chen, Hui-Chen Lu, Raymond A Chitwood & Daniel Johnston
ダニエル・ジョンストン。
Neural correlates of binocular rivalry in the human lateral geniculate nucleus
Klaus Wunderlich, Keith A Schneider & Sabine Kastner
Neural basis and recovery of spatial attention deficits in spatial neglect
Maurizio Corbetta, Michelle J Kincade, Chris Lewis, Abraham Z Snyder & Ayelet Sapir
Perceptions of moral character modulate the neural systems of reward during the trust game
M R Delgado, R H Frank & E A Phelps
上の3本は未読。あとで読みます。
ところで、僕にとって、最新号の論文誌をチェックをすることの目的のひとつは、専門外だけど興味ある分野(たとえば認知・心理系など)に定期的に目を通すこと。それによって、目を広げれたらいいな、と。
でも結局、いそがしいと専門分野の論文だけ目を通して、専門外の論文は後回しにしちゃうから、目的が達成されてませんね。。
投稿者 sfujisawa : 20:51 | コメント (0)
2005年10月28日
Phys Rev Lett 10/28
Entangled Networks, Synchronization, and Optimal Network Topology
Luca Donetti, Pablo I. Hurtado, and Miguel A. Munoz
「Entangled network」という新たなネットワークtopologyを見つけたよー、という論文。
最近はやりの、Network に関する研究(たとえば Small-world network など)は、おもに topology に焦点をあてたものが多い。確かに、人間関係のネットワークやWWWでのネットワークの研究では、topologyのみに注目してても、十分面白い。しかし、Neural networkでは、topologyに加えてdynamicsも知りたい。あるtopologyをもつネットワークにおいて、各ノード(ニューロン)が時々刻々とある活動をしているときにネットワーク全体としてどういうdynamicsになるか、ということに興味があるわけである。
ところが、BarabasiにしろWattsにしろNewmanにしろ、この手の研究で有名な人たちは、dynamicsにはあまり興味がないようです。(Barabasiなんか、scale-free network の結合行列の固有値まで求めているくせに、dynamicsには目も向けてない。。)
今回の論文は、topologyとdynamicsの両方に注目した論文です。
--
Pecoraの論文(PRL1998やPRE1998など)では、ネットワークのsynchronizationの安定性に関する議論をしています;
ひとつの振動子のdynamicsは、
dx / dt = F(x)
で記述できるとします。XはベクトルでもOKです。(Pecora論文では、x を3次元ベクトルとして Rossler Attractor などを考えたりしています。)この振動子が、N個存在して、お互いがinteractしているとき、i 番目の振動子の微分方程式を、
dxi / dt = F(xi) + σ Σj Lij H(xj)
と記述します。ここで、L は結合行列で、Lii = -k (kは、ノード i が有する結合数) 、Lij = 1 (ノード i と j が結合があるとき)、Lij = 0(ないとき)とします。(つまり、L は対称行列。)ここから、すべてのノードがsynchronizeしているときからの摂動を評価することにより、synchronizationの安定性を評価する、という流れみたいです。(途中はあんまりよく理解できなかった。)
結局、行列 L の固有値をもとめて、
固有値比 Q = (最大固有値) / (0以外の最小固有値)
が小さいほど安定性が高い、ということらしいです。トポロジーによって決まる結合行列 L から synchronization dynamics の安定性が議論できる、というのがミソ。
それでは、ネットワークのノード数 N と、平均結合数
まずはじめに、例として、small-world network や random network や linear chainなどの Q を計算している。(この中では、small-world の Q がいちばん小さいらしい。)つぎに、Q が小さくなるように、これらのネットワークから(あるアルゴリズムによって)ノードをひとつひとつつなぎ変えていく。そうしてsその極限で得られたのが、この論文のFig1の右側のようなネットワーク。このネットワークの特徴は、
・各nodeの性質が非常に似通っている。
・コネクションはとても複雑で入り組んでいる。
・Modularityは乏しい。
など。このネットワークは、“ Entangled network ” (もつれネットワーク?)と名付けられてます。
実際、神経細胞のネットワーク(たとえばCA3のリカレントネットワーク)を考えてみると、small-worldよりもこっちのentangled networkの方が近いかもね。なんとなくですけど。
まあもっとも、この論文やPecora論文で言うところのsynchronizationと、神経生理学で興味のあるsynchronizationとではまた意味が違うのですが、まあでもnetworkのdynamicsに焦点をあてている論文、ということで長々と書いてみました。
投稿者 sfujisawa : 20:21 | コメント (0)
2005年10月20日
Neuron 10/19
Progressive Transformation of Hippocampal Neuronal Representations in ""Morphed"" Environments
J.K. Leutgeb, S. Leutgeb, A. Treves, R. Meyer, C.A. Barnes, B.L. McNaughton, M.-B. Moser, and E.I. Moser
Moser、またかぃ!
ラット海馬CA1とCA3での place cell についての論文。実験のパラダイムは、前回のO'Keefeの論文(Wills et al Science 2005)(参考)とほぼ同じ。円形の実験箱と四角形の実験箱を使う。Place cell は円形箱と四角形箱では、place field が異なる(そうなるまでプレトレーニングで学習させる)。このとき、実験箱を、円形から四角形に徐々に(7段階ぐらいで)変形していったら、place cellはどう発火するか、という実験。
O'Keefeの実験では、丸い空間を徐々に四角い空間に変えていったら、place cell の place field は、ある時点で、不連続に、丸い空間の place field から四角い空間の place field へ遷移するという結果が得られている。今回のMoser論文では、O'Keefe論文のようにplace fieldが不連続に遷移するという細胞(Figure3、"Sigmoid")と、連続的に段階的に変化するという細胞("Linear")の両方が存在したという結果(CA1、CA3ともに)。
CA1とCA3での違いについては、CA3の place cell ほうが、CA1のよりも、異なった環境(円形箱と四角形箱)をより分離する傾向がある、ということらしい。(Place field の overlap率のデータなどから。)
さて、最近似たような論文がでてこんがらがってきたので、明日ぐらいまたちょっとまとめてみます。
投稿者 sfujisawa : 22:25 | コメント (0)
2005年10月14日
Science 10/14
Observing Others: Multiple Action Representation in the Frontal Lobe
Koen Nelissen, Giuseppe Luppino, Wim Vanduffel, Giacomo Rizzolatti, and Guy A Orban
サル、fMRI。ミラーニューロンネタ。(勉強しなければ。。)
投稿者 sfujisawa : 22:51 | コメント (0)
2005年10月12日
Nature 10/12
メモ。
Repeated cocaine exposure in vivo facilitates LTP induction in midbrain dopamine neurons
Qing-song Liu, Lu Puand Mu-ming Poo
Pooは最近こんなことやってんですね。コカイン投与はin vivo(腹腔)ですが、電気生理はスライスみたいです。
投稿者 sfujisawa : 21:24 | コメント (0)
2005年10月07日
Science 10/7
ところで、来年のSfNの annual meeting は、アトランタに変更になったみたいですね。
Failure to Detect Mismatches Between Intention and Outcome in a Simple Decision Task
Petter Johansson, Lars Hall, Sverker Sikström, and Andreas Olsson
以下、先週のScience。読むの忘れてた。
Breakdown of Cortical Effective Connectivity During Sleep
Marcello Massimini, Fabio Ferrarelli, Reto Huber, Steve K. Esser, Harpreet Singh, and Giulio Tononi
ヒト、TMS(経頭蓋的磁気刺激法)とEEG(60電極)。覚醒時とNonREM睡眠時で、皮質にTMS刺激を与えたときに、TMS-evoked response をEEGで調べて比較した、という内容。覚醒時では、TMS-evoked response は 300ms ほど続き、かつ数センチ離れた皮質まで伝播する。しかし、NREM時では、TMS-evoked responseのamplitudeは覚醒時より大きいものの、120msほどで減衰し、また伝播も観察されなかった。このような覚醒時と睡眠時における皮質の「effective connectivity」の違いを、consciousnessと絡めて議論しています。
Direct Evidence for a Parietal-Frontal Pathway Subserving Spatial Awareness in Humans
Michel Thiebaut de Schotten, Marika Urbanski, Hugues Duffau, Emmanuelle Volle, Richard Levy, Bruno Dubois, and Paolo Bartolomeo
ヒト、Intraoperative electrical stimulationという手法。これは、Glioma(神経膠腫)患者の腫瘍摘出手術中に、摘出部位の境界を決定するために行うものらしい。頭蓋を開けた状態で、脳表の摘出予定部位に直接電極をあて、電気刺激をしてその部位の活動を一時的に抑制する。そのとき、患者が一時的に重要な障害を生じれば(たとえば会話ができなくなるとか)、その部位は摘出するのを止める、というような感じで。
ここで、(右半球の)caudal superior temporal gyrus (cSTG) および supramarginal gyrus (SMG)を電気刺激したとき、線分を二等分する課題で、障害を生じた(分離点が右にずれた)、という結果。(なので、この部位の摘出は止めた。)この部位は、Parietal-Frontal pathwayに相当する部位らしいが、このpathwayが視覚のsynmetrical processingに必要である、という結論。
この論文では特に半側性空間無視などと絡めて議論しているが、この辺は勉強不足なので、pooneilさんのサイトを復習して勉強中です。
投稿者 sfujisawa : 21:52 | コメント (0)
2005年10月06日
Neuron 10/5
メモ。
Perceptual Organization of Tone Sequences in the Auditory Cortex of Awake Macaques
Christophe Micheyl, Biao Tian, Robert P. Carlyon, and Josef P. Rauschecker
サルの電気生理&ヒト。A、Bの別の周波数をもつ音を、A-B-A A-B-A ・・・と繰り返し発生させるとする。AとBの周波数の差ΔFが小さければ、「A-B-A」と聞こえるし、ΔFが大きければ、「A-A-A-A」「B-B」と別の2つのsteamとして聞こえる。この現象に対して、ヒトでの心理物理学的実験と、サル一次聴覚野でのユニット記録の実験を行ったもの。
Updating Visual Space during Motion in Depth
Nuo Li and Dora E. Angelaki
投稿者 sfujisawa : 20:54 | コメント (0)
2005年10月05日
JNS 10/5
Scopolamine Reduces Persistent Activity Related to Long-Term Encoding in the Parahippocampal Gyrus during Delayed Matching in Humans
Karin Schon, Alireza Atri, Michael E. Hasselmo, Marisa D. Tricarico, Matthew L. LoPresti, and Chantal E. Stern
ヒト、fMRI。(1)視覚のdelayed matching-to-sample (DMS) taskを行っているとき、delayの時のparahippocampusの活動をfMRIで測定。(これを、persistent activityと解釈している。)スコポラミン(ムスカリンR-アンタゴ)を投与された被験者では、このpersistent activityは減少する。(2)DMSの20分後に、DMSで使った画像を覚えているかのtask(これをlong-term momoryとしている)。スコポラミン群ではこのスコアも減少していた。つまり、DMSでのdelay時のpersistent activityと長期記憶の関係(相関)と、そのときのコリン系の役割を調べた研究。
NMDA/AMPA Ratio Impacts State Transitions and Entrainment to Oscillations in a Computational Model of the Nucleus Accumbens Medium Spiny Projection Neuron
John A. Wolf, Jason T. Moyer, Maciej T. Lazarewicz, Diego Contreras, Marianne Benoit-Marand, Patricio O'Donnell, and Leif H. Finkel
コンピューターモデル実験。ここでのstate transitionは、UP/DOWNでのDOWN←→UPのtransitionのことでした。
Behavioral Evidence That Segregation and Representation Are Dissociable Hippocampal Functions
Stepan Kubik and Andre A. Fenton
ラット、行動課題。あまりよくわからなかった。
Hormonal Cycle Modulates Arousal Circuitry in Women Using Functional Magnetic Resonance Imaging
Jill M. Goldstein, Matthew Jerram, Russell Poldrack, Todd Ahern, David N. Kennedy, Larry J. Seidman, and Nikos Makris
投稿者 sfujisawa : 20:42 | コメント (0)
2005年10月04日
Nature Neurosci 10月号
時差ぼけで10時睡眠-4時起床の生活(ある意味健康的)。げりもまだなおらん。
NNの最新号を読み忘れていたので、軽く目を通してメモ。
Dynamics of excitation and inhibition underlying stimulus selectivity in rat somatosensory cortex
W Bryan Wilent & Diego Contreras
ラットの in vivo イントラ記録。Somatosonsory cortex の thalamoreceipient neuron から。このニューロンでは、ひげを動かす方向に対して方向選択性があるらしい。その方向選択性のメカニズムについて、興奮性入力と抑制性入力それそれのコンダクタンスを詳細に測定することで明らかにしようとした論文。
Asynchronous GABA release generates long-lasting inhibition at a hippocampal interneuron principal neuron synapse
Stefan Hefft & Peter Jonas
海馬 DG で、interneuron と granule cell をダブルパッチして、interneuronの電気生理学的性質を調べたもの。この論文では、CCK-positiveなinterneuronでは、強い発火が生じた後では、somaの発火タイミングに依存しないGABAの放出(←つまりminiatureみたいなもの?)が存在する、という発見。これを、asynchronous GABA releaseと呼んでいる。PV-positiveなinterneuronではこの現象は無いらしい。
Cortical responses to invisible objects in the human dorsal and ventral pathway
Fang Fang & Sheng He
ヒト、fMRI。左右の目に別々の画像を見せられるようにした状態で(つまり視野闘争が生じている状態で)、dominantな方の目にはノイズ画像を見せて、non-dominantの方の目にはobject画像を見せる。(つまり、意識下ではobject画像は認識されていない。)このとき、dorsal pathwayとventral pathwayでどう反応が異なるか、を調べたもの。結果は、dorsalでは、このようなときも、objectが認識されていると時と同様に強く反応するというもの。しかも、object画像が、人間の顔の画像の時よりも、道具の画像の時の方が強く反応するとか。(これは、dorsal pathway(つまり"where" pathway)が、reachingやgraspingに重要な役割を持っているからではないかと考察している。)
Tuning for spectro-temporal modulations as a mechanism for auditory discrimination of natural sounds
Sarah M N Woolley, Thane E Fremouw, Anne Hsu & Frederic E Theunissen
鳥(zebra finch)、ユニット記録。(鳥はちょっと苦手。)
Timing of the brain events underlying access to consciousness during the attentional blink
Claire Sergent, Sylvain Baillet & Stanislas Dehaene
投稿者 sfujisawa : 20:56 | コメント (0)
2005年09月16日
Neuron 9/6
今日は手抜きでメモのみ。
Roles of NMDA NR2B Subtype Receptor in Prefrontal Long-Term Potentiation and Contextual Fear Memory
M.-G. Zhao, H. Toyoda, Y.-S. Lee, L.-J. Wu, S.W. Ko, X.-H. Zhang, Y. Jia, F. Shum, H. Xu, B.-M. Li, B.-K. Kaang, and M. Zhuo
海馬ではNR2AがLTP、NR2BがLTDという報告が去年あったが、PFCではNR2BもLTPに関わるよー、という論文。
Dissociable Controlled Retrieval and Generalized Selection Mechanisms in Ventrolateral Prefrontal Cortex
D. Badre, R.A. Poldrack, E.J. Pare-Blagoev, R.Z. Insler, and A.D. Wagner
Wagnerラボ。ヒト、fMRI。4種類の言葉の選択課題(judgment specificity, cue-target-associative strength, competitor dominance, and number of competitors:論文Fig1参照)を行う。このとき、ventrolateral prefrontal cortex (VLPFC)の各部位で、課題の種類によって活動レベルが異なった、という話し。
Dynamics of Parietal Neural Activity during Spatial Cognitive Processing
D.A. Crowe, B.B. Averbeck, M.V. Chafee, and A.P. Georgopoulos
GABAergic Excitation Promotes Neuronal Differentiation in Adult Hippocampal Progenitor Cells
Y. Tozuka, S. Fukuda, T. Namba, T. Seki, and T. Hisatsune
投稿者 sfujisawa : 21:45 | コメント (0)
2005年09月13日
PNAS 9/13
Orthogonal arrangement of rhythm-generating microcircuits in the hippocampus
Tengis Gloveli, Tamar Dugladze, Horacio G. Rotstein, Roger D. Traub, Hannah Monyer, Uwe Heinemann, Miles A. Whittington, and Nancy J. Kopell (Contributed by Kopell)
海馬スライスの電気生理。3次元的な構造をもつ海馬を、いろいろな面でスライスにして、カイニン酸をかけたときのgamma/thetaの発生・同期がどう異なるかを評価。Networkの方向性に注目した論文。スライスの作り方は以下の3種類;
・transvers(Ventralの海馬をholizontalに切った平面。いちばん一般的な平面)
・longitudinal(Ventralの海馬をsagittalに切った平面)
・coronal(Ventralの海馬をcoronalに切った平面)
transversスライスでは、gammaは発生するが、thetaは発生しない、longitudinalスライスでは、thetaは発生するが、gammaは発生しない。coronalスライスでは、両方発生する、という結果。組織染色もやっていて、O-LM cellの投射方向の違いなどに注目してdiscussionしています。
投稿者 sfujisawa : 19:59 | コメント (0)
2005年09月08日
JNS 9/7
メモのみ。
Retrieval Attempts Transiently Interfere with Concurrent Encoding of Episodic Memories But Not Vice Versa
Kevin Allan and Roy Allen
ヒト、行動試験とEvent-related potential。StageIでは、音声により単語("mother"とか"rock"とか)を与えられてそれが生き物であるかを判定する課題(encode)。StageIIでは音声により単語を与えられてそれがStageIで出てきた単語かを判定する課題(retrieval)と、視覚により単語を与えられてそれが生き物であるかを判定する課題(encode)を、音と視覚のタイミングを少しだけずらして同時にやる(つまり、encodeとretrivalの同時課題)。StageIIIは、視覚により単語を与えられて、それがStageIIで出てきた単語か判定(retrieval)。それで、encodeとretrievalの同時課題では、retrievalは変化ないが、encodeは影響されてスコアが下がる、という内容。
Amygdalar and Prefrontal Pathways to the Lateral Hypothalamus Are Activated by a Learned Cue That Stimulates Eating
Gorica D. Petrovich, Peter C. Holland, and Michela Gallagher
ラット、immediate-early gene系の論文。
Multiple Object Response Normalization in Monkey Inferotemporal Cortex
Davide Zoccolan, David D. Cox, and James J. DiCarlo
投稿者 sfujisawa : 20:53 | コメント (0)
2005年09月02日
Neuron 9/1
今日は手抜きでメモのみ。
Memory Strength and Repetition Suppression: Multimodal Imaging of Medial Temporal Cortical Contributions to Recognition
Brian D. Gonsalves, Itamar Kahn, Tim Curran, Kenneth A. Norman, and Anthony D. Wagner
Wagnerラボ。
Adaptation to Synaptic Inactivity in Hippocampal Neurons
T.C. Thiagarajan, M. Lindskog, and R.W. Tsien
投稿者 sfujisawa : 22:26 | コメント (0)
2005年09月01日
JNS 8/31
Self-Motion and the Hippocampal Spatial Metric
Alejandro Terrazas, Michael Krause, Peter Lipa, Katalin M. Gothard, Carol A. Barnes, and Bruce L. McNaughton
McNaughtonラボ。ラット、海馬CA1のユニット記録。ラットが、同じ環境下において、自分で歩いているときと、車に乗って移動しているとき(!)ではplace cellの発火パターンは変化するのか、という実験。車の時は、Theta波自体が小さくなり、またTheta波に対するpryramidal cellの発火phaseのlockが少し甘くなり、phase precessionも甘くなる、という結果。
これ、実験風景のビデオをsupplementalにつけてくれると面白かったのに。。ちなみに、車はこれを改良して使ったらしい。
投稿者 sfujisawa : 22:05 | コメント (2)
2005年08月31日
Nature Neurosci 9月号(3)
最近とくにネタがないので、今日も文献の紹介です。。
Risk-sensitive neurons in macaque posterior cingulate cortex
Allison N McCoy & Michael L Platt
サル、postrior cingulate cortex(CGp,後帯状回?)でのユニット記録。(CGpについては僕はよく知らないのですが。)CGpにはリスクに反応するニューロンがある、という話し。Taskは、visualギャンブル課題。ターゲット(visual)が二つあって、片方のターゲットでは常に報酬は同じであるが、もう片方は報酬がギャンブル的(平均すると同じ)で、サルはどちらかを(凝視することによって)選ぶ。CGpニューロンの視覚受容野内に存在するターゲットが'risky'targetであり、そのtargetをchoiceしたとき('risky'choiceをしたとき)に、このニューロンの発火率が上昇する、という結果。(そのニューロンの受容野外のターゲットにたいして'risky'choiceをした場合は、発火率に変化はない。)
投稿者 sfujisawa : 21:47 | コメント (0)
2005年08月30日
Nature Neurosci 9月号(2)
Neural codes for perceptual discrimination in primary
somatosensory cortex
Rogelio Luna, Adrian Hernandez, Carlos D Brody & Ranulfo Romo
Romo のグループ。彼らの、vibration刺激のfrequency descrimination taskを用いた一連の研究(RomoのNRNとか参考)の続報。サルの指にバイブレーターを当てて、まず周波数f1の振動刺激をあたえて、少し時間をおいて(3s)、次に周波数f2の振動刺激を与えて、f1とf2の振動のどちらの周波数が高かったかを判断させる課題。このとき、primary somatosensory cortexでは、刺激の周波数にロックして発火する細胞が観察されるが(RomoのPNAS200とか)、ここでの発火が、どのようにコードされて、最終的なdescriminationに用いられるか、というところに焦点をあてた、データ解析論文。ISI、平均発火率、発火数、バースト率、バースト数などの量を解析してみたが、結局、250msのtime windowで区切ったときの総発火数の曲線が、もっとも行動曲線に近い、という結論。つまり、一次体性感覚野での250ms程度の時間区間での発火数の和が、(より高次の皮質で)descriminationの判断に使われるというのが彼らの推論。
Neural basis of auditory-induced shifts in visual time-order perception
John J McDonald, Wolfgang A Teder-Salejarvi, Francesco Di Russo & Steven A Hillyard
ヒト、event-ralated potentials。左右に電球がひとつずつあって、どちらが先に光ったかを判定するtask。ここで、それぞれの電球の下にはスピーカーがセットしてある。電球が光るまえに、どちらかのスピーカー(たとえば右スピーカー)の音を鳴らした場合、左右の電球が同時に光ったとしても、ヒトは右の電球が先に光ったと感じるらしい。これを「auditory-induced shifts in visual time-order perception」といっているのだが、このときのERPを測定した実験。音刺激が存在したとき、後頭部でのERPのピークの時間は変わらないが、ピークの大きさが変化する、という結果。
投稿者 sfujisawa : 22:05 | コメント (0)
2005年08月29日
Nature Neurosci 9月号
今月号はやけに認知・生理系の論文が多いなぁ。少しずつ読んでみます。
Auditory thalamus integrates visual inputs into behavioral gains
Yutaka Komura, Ryoi Tamura, Teruko Uwano, Hisao Nishijo and Taketoshi Ono
小野武年先生のグループ。ラット・Auditory thalamusのユニット記録。Taskはauditory spatial discrimination task。Cue として、音 cue を用いるもの、光 cue を用いるもの、音+光 cueを用いるものを行う。音 cue を用いた課題(unimodal)の場合、auditory thalamusは音に反応する。光 cue を用いた課題(unimodal)では、auditory thalamusはほとんど反応すしない。しかし、音+光 cue を用いた課題(bimodal)では、単に音だけの時に比べて発火率が約2倍になる、という結果。つまり、光入力は、auditory thalamusにたいして、gain modulationを行う、というもの。あと、Rewardも絡めています。
投稿者 sfujisawa : 20:41 | コメント (0)
2005年08月24日
Nature 8/25
The neuronal representation of pitch in primate auditory cortex
Daniel Bendor and Xiaoqin Wang
Pitch(「音程」と訳せばいいのかな?)に反応するニューロンを見つけた、という話し。サル、電気生理(ユニット記録)。このニューロンは基調周波数f0のharmonicに反応するらしい。このようなニューロンは、primary auditory cortexのantero-lateral側の近傍に存在するとか。
投稿者 sfujisawa : 22:36 | コメント (0)
2005年08月17日
Neuron 8/18
軽くメモだけ。
Brain Mechanisms for Extracting Spatial Information from Smell
J. Porter, T. Anand, B. Johnson, R.M. Khan, and N. Sobel
ヒトでのfMRI実験。人間や動物ははある匂いを嗅いだときに、匂い源をだいたい特定することができる。匂い源の場所特定をしているときのprimay olfactory cortexやsuperior temporal gyrusのfMRIイメージングを測定したというもの。
Differential Neural Responses Evoked by Orthonasal versus Retronasal Odorant Perception in Humans
D.M. Small, J.C. Gerber, Y.E. Mak, and T. Hummel
こちらも匂いネタ。鼻経路で匂いが入ってきたときと口腔経路で匂いがはいってきたときの違いをfMRIイメージングしてる。
投稿者 sfujisawa : 22:39 | コメント (0)
2005年08月11日
JNS 8/10
今週号の表紙の Growth cone はきれいですね。
Prefrontal Control of the Amygdala
Ekaterina Likhtik, Joe Guillaume Pelletier, Rony Paz, and Denis Pare
Pare研。Prefrontal cortex → BLA のprojectionの電気生理学的研究。Pare研はうちと同じ研究棟にあって、ちょうどうちらの下の階です。よくPareがぶらぶら歩いているのを見かけます。
投稿者 sfujisawa : 18:38 | コメント (0)
2005年08月10日
Nature 8/11
Microstructure of a spatial map in the entorhinal cortex
Torkel Hafting, Marianne Fyhn, Sturla Molden, May-Britt Moser and Edvard I. Moser
Moser&Moserのグループ。Entorhinal cortexではグリッド状に空間情報をコードするplace cellが存在する、という内容。これもうちでは早くから流れてた。
投稿者 sfujisawa : 20:37 | コメント (0)
2005年08月04日
JNS 8/3
今日は手抜きでメモだけ。すみません。。
Short-Term Depression in Thalamocortical Synapses of Cat Primary Visual Cortex
C. Elizabeth Boudreau and David Ferster
ひさびさのFersterラボ。
The Human Hippocampus: Cognitive Maps or Relational Memory?
Dharshan Kumaran and Eleanor A. Maguire
Encoding and the Durability of Episodic Memory: A Functional Magnetic Resonance Imaging Study
Melina R. Uncapher and Michael D. Rugg
投稿者 sfujisawa : 21:53 | コメント (0)
2005年08月03日
Neuron 8/4
Inhibitory Postsynaptic Potentials Carry Synchronized Frequency Information in Active Cortical Networks
A. Hasenstaub, Y. Shu, B. Haider, U. Kraushaar, A. Duque, and D.A. McCormick
McCormickのグループ。スライスとvivoの両方でUP/DOWN中でのEPSP入力とIPSP入力を詳細に調べた、という論文。UP中ではIPSP入力は近隣細胞で同期してるよ、だからIPSPは発火のシンクロやタイミングモジュレーションに重要だよ、という内容。それほど真新しくはない。(というか、なんかマンネリ化してきたなー。。)
Synaptic Mechanisms of Forward Suppression in Rat Auditory Cortex
M. Wehr and A.M. Zador
感覚刺激、例えば音刺激を2回連続(刺激間隔 ~500ms)で与えると、auditory cortexのneuronの発火数は、1回目の刺激に比べて、2回目の刺激では減少する。これをforward supression(あるいは forward masking)というらしい。このメカニズムを調べた論文。Vivoパッチで、音刺激を与えたときのPSPとそのときのSynapticコンダクタンスを評価した実験。Synapticコンダクタンスは、Excitatory成分もInhibitory成分も50~100msで元に戻るので、100ms以上持続するforward supressionは、GABA系による抑制が原因ではない、という結論。
Zador、久しぶりだなー、と思って調べてみたら、この論文を見落としてた。最近はvivoパッチでコンダクタンスを調べる系のことをやっているのですね。
Testing the Efficiency of Sensory Coding with Optimal Stimulus Ensembles
C.K. Machens, T. Gollisch, O. Kolesnikova, and A.V.M. Herz
面白そうなのですが、あまりよく分かりませんでした。
投稿者 sfujisawa : 21:32 | コメント (0)
2005年07月29日
Phys Rev Lett 7/29
Phase Transition in a Healthy Human Heart Rate
Ken Kiyono, Zbigniew R. Struzik, Naoko Aoyagi, Fumiharu Togo, and Yoshiharu Yamamoto
人間の心臓の拍動を記録して、それが phase transition していることを示した論文。なんか、fluctuation の correlation を計算するような解析で、心臓の拍動が"second order phase transition"だと言っている。心臓の拍動のような単純なものでもphase transition (like)と決定できるような解析法ならけっこう使えそうだとは思うのだが、難しくて何をやっているのかよくわからん。。あとでゆっくり読もうか。
投稿者 sfujisawa : 22:26 | コメント (0)
2005年07月28日
Nature 7/28
Robust habit learning in the absence of awareness and independent of the medial temporal lobe
Peter J. Bayley, Jennifer C. Frascino and Larry R. Squire
Squireのグループ。例によって側頭葉損傷患者の実験。ヒトにおいて、object recognition taskが、declarative的にだけでなく、Habit的にも記憶されうる(ただし側頭葉非依存)ことを示した論文。(サルでは知られてたらしい。)
Task:Objectは、変な形のプラスチックや金属の破片など。それぞれが紙のカードの上にひっつけられている。Objectカードは16枚あって、2枚ずつ8組のpairになっている。それぞれのpairで、"correct" objectが一枚決められていて、そのobjectカードの裏側に"correct"の文字が書かれている。
被験者は、1pairずつ提示されて、「この2枚のうち、"correct"objectを、try-and-error(つまり自由にobjectカードをつまんでひっくり返して"correct"の文字があるかどうか確かめられる)で覚えてください」と指示される。1セッションは40trial、週に2回のセッションを行う。(The concurrent discrimination task)
Controlでは、だいたい3回のセッションで、90%以上の正解率。
ここで側頭葉損傷患者(EPさんとGPさん)では30回程度のセッションで85~90%程度の正解率でこのobject recognition taskが学習された。しかし、彼らはこのtrialをしたこと自体は覚えていないし、このtaskがどういうものなのかの説明もできない。(つまり「declarative memory」ではない。)
次に、記憶が形成された後に、16枚のカードを同時に並べて、「この中から"correct"objectを選んでください」と指示される課題(The sortig task)。Controlはできるが、側頭葉損傷患者は全くできない。しかし、この直後に、1pairずつ提示されるtask(今度はつまむのではなく言葉で(右か左か)答えなくてはならない)は、答えられる。
さて、Taskを行ったこと自体は覚えていないけど、Habit memoryは獲得される(たとえば鏡を見ながら鉛筆で図形をトレースするtaskとか)ことは、海馬損傷患者の研究(たとえばHMさんとか)で相当前から知られている。
今回新しいのは、Object recognition taskのような「declarative memory」と考えられていたものも、Habit learningで獲得できることを示したことかな?あるいは、Object recognition のようなtaskにおいて、海馬・側頭葉経路で意識下にdeclarative的にすばやく記憶される経路と、basal ganglia経路(?)で無意識下にHabit的にゆっくり記憶される経路の2種類が存在することを示したことかな。
投稿者 sfujisawa : 21:01 | コメント (2)
2005年07月25日
Science 7/22
Independent Codes for Spatial and Episodic Memory in Hippocampal Neuronal Ensembles
Stefan Leutgeb, Jill K. Leutgeb, Carol A. Barnes, Edvard I. Moser, Bruce L. McNaughton, and May-Britt Moser
Moser&Moser&McNaughton。Moser妻がLast autherで、McNaughtonがコレスポという微妙な論文。CA1/CA3での place cellの論文。大きい環境(実験室)とLocalな環境(実験箱)をplace cellはどうコードしているかという内容。まず、同じ実験箱でも、実験室が変わると、place cell のコードする領域が変わるのは以前から知られていた(Figure2)。(つまり、place cellは distal な cueを重要とする。)さて、今回の新しい発見は、実験室は同じで、実験箱を変えた場合、一見、place cellのコードする領域が変わったように見えるが、実は発火数が変わっているだけで、コードしている領域は同じだ、というもの(Figure1)。つまり、localのcueは、発火率の違いによりコードしている、という解釈。
タイトルに「Episodic memory」とあるが、内容は episodic memoryとは全く関係ない。
ところで、この論文のPreviewはギューリーが書いている。「ジャガーに乗っているときとロータスに乗っているときでは、同じ道を走っていてもplace cellは違うふうに発火するよーん」ってたとえてます。(つまり車がlacal cueで、道がdistal cueってわけね。)
この論文とPreviewの草稿はうちでは早くから流れていてセミナーでもやったが、べつにギューリーが論文のレビューをしたわけではないらしい。ちなみにジャガーとロータスはギューリーの持ってる車で、草稿では自分の車の写真をFigureに載っけてたのだが、publishされたFigureでは車の写真がイラストになってたから、science編集部に直されたんだねー。あと、草稿では、「"Episodic memory"はタイトルに書いてるだけで本文でふれてないやん」って皮肉ってたけど、これも消されてた。。
投稿者 sfujisawa : 21:36 | コメント (0)
2005年07月21日
Neuron 7/21
The Coordinated Mapping of Visual Space and Response Features in Visual Cortex
H. Yu, B.J. Farley, D.Z. Jin, and M. Sur
Sur研。一次視覚野の simple cell/complex cell は、網膜での位置(2次元)、方位選択性(2次元)、眼優位性(ocular dominance、1次元)などに対して反応する。つまり、入力される情報は5次元である。で、一次視覚野では2次元空間的にこれらの情報をmappingしているので、demension reductionが生じている。ここで、入力の直交性を保ったまま(つまり各次元の独立性が保たれたまま)demension reductionしているかどうかは興味深い問題。この論文では、一次視覚野の応答の精密な測定と、理論とでその問題を研究している。でもSurの論文は難しくてよく分からん。
投稿者 sfujisawa : 22:01 | コメント (0)
2005年07月19日
PNAS 7/19
Synaptic mechanisms of persistent reverberatory activity in neuronal
networks
Pak-Ming Lau and Guo-Qiang Bi(Edited by CF Stevens)
Bi研。Networkのstimulus-induced persistent activityを調べた論文。 実験系はCultureのシングル・パッチ。特に目新しいところはなかった。Buonomano の2003 PNASの方が上かな。
投稿者 sfujisawa : 22:12 | コメント (0)
2005年07月15日
Science 7/15
Evidence for Ectopic Neurotransmission at a Neuronal Synapse
JS Coggan, TM Bartol, E Esquenazi, JR Stiles, S Lamont, ME Martone, DK Berg, MH Ellisman, TJ Sejnowski
なぜこのタイトルでSejnowski?とおもったら、コンピューターモデル実験だった。電顕写真から3次元再構築したモデルシナプスをつかってモンテカルロシミュレーションを行い、synaptic releaseされた ACh が extra-synapticなレセプターを活性しうることを示したもの。Sejnowski、恐るべし。
投稿者 sfujisawa : 19:18 | コメント (0)
2005年07月07日
Nature 7/7
Dynamic predictive coding by the retina
Toshihiko Hosoya, Stephen A. Baccus and Markus Meister
Meisterのグループ。いつもの、salamander(サンショウウオ)の網膜を組織切片として単離して61電極arrayにのせてganglion cellの発火のunitを調べる実験。網膜では、視覚入力(光)に対して、入力の絶対値をコードしているのではなく、入力の変化をコードしているらしい。では、動物が異なる環境下に移動した場合、網膜はどのようにadaptaionを行っているか、というところに焦点をあてた論文。難しそうなのであまり詳しく読んでない。
Nonlinear dynamics: When instability makes sense
Peter Ashwin and Marc Timme
News&Viewsなんだけど、紹介している論文は古い論文ばっかりだった。。論点は、力学系ダイナミクスの安定性について。力学系で議論されるのは、安定点や安定なリミットサイクルのことが多いが、ニューラルネットワークなどのダイナミクスでは、不安定な鞍点が重要なのではないか、という提案。つまり、リミットサイクルでは安定な状態に落ち込んでしまっている状態だが、鞍点は不安定でその場に長い間とどまることができないので鞍点間の飛び移りが生じるので、sequence情報をencodeするのに重要なのではないか、という話し。それで、紹介されていたのが以下の論文↓:
Reproducible sequence generation in random neural emsembles
Huerta R and Rabinovish M, Phys Rev Lett 93,238104 (2004)
興奮性(E)、抑制性(I)を含むランダム結合のニューラルネットで、どのような結合の時に、繰り返し生じる「Sequence」や、リミットサイクルが生じるか、ということを調べた論文。リミットサイクルは、E-Iバランスがとれているときに生じやすいが、Reproducible sequenceはEIバランスには関係ない、という結論。(Cortical Songを彷彿とさせる論文ね。)ただ、力学系の問題などがきちんと議論されているわけではなく、PhysRevLettにしてはなにか不完全燃焼的な印象を受ける論文。
これ系の内容は、neural network の sequence code の問題に対して本質的に重要だから、きちんと数学的に議論できる枠組みが早くできたらいいのにな。
投稿者 sfujisawa : 21:41 | コメント (0)
2005年07月01日
JP 7/1
Dynamic synapses as archives of synaptic history: state-dependent redistribution of synaptic efficacy in the rat hippocampal CA1
Takuya Yasui, Shigeyoshi Fujisawa, Masako Tsukamoto, Norio Matsuki, and Yuji Ikegaya
ヤスの論文。僕も解析の手伝い(Mutual Entropyのところ)で名前を入れてもらいました。それにしても、彼はよく頑張ったなー。17ページの大論文ですね。
投稿者 sfujisawa : 20:10 | コメント (0)
2005年06月30日
JNS 6/29
A Subpopulation of Neurons in the Medial Prefrontal Cortex Encodes Emotional Learning with Burst and Frequency Codes through a Dopamine D4 Receptor-Dependent Basolateral Amygdala Input
Steven R. Laviolette, Witold J. Lipski, and Anthony A. Grace
AA Graceのグループ。恐怖条件付け課題を行ったときの、mPFCとBLAのinteractionが存在するということ(CSを与えることにより、反応するmPFCニューロンが存在し、BLAにmuscimolを注入してBLAの機能を抑制するとそのmPFCニューロンの反応もなくなる、という現象)と、それが D4 recepter dependentである(mPFCにD4アンタゴを打ち込むと反応が消える)ことを示した論文。それにしても、麻酔下でも恐怖条件付けができる、っていうのはおもしろいですね。彼らのこの論文とかはけっこう好き。
投稿者 sfujisawa : 21:46 | コメント (0)
2005年06月24日
Phys Rev Lett 6/24
Anomalous Transport in Scale-Free Networks
Eduardo Lopez, Sergey V. Buldyrev, Shlomo Havlin, and H. Eugene Stanley
Stanleyのグループ。ネットワークのノード間の"conductance"を調べるというもの。(各ノードが単位resistanceを持っているとして。)ネットワーク内のTransportに焦点を当てた研究。よく読んでない。
投稿者 sfujisawa : 22:21 | コメント (0)
2005年06月23日
Nature 6/23
Invariant visual representation by single neurons in the human brain
R. Quian Quiroga, L. Reddy, G. Kreiman, C. Koch and I. Fried
人間の海馬の単一細胞からのユニット記録(!!)。重度の側頭葉てんかんの患者さんに、治療目的で電極を埋め込むことはけっこうやられていて、患者にコンセンサスをとってその電極から研究目的の記録をおこなうという論文はたまに見かけていた。しかし、ユニット記録を行うとなると、電極は4本以上埋め込まないといけない。この論文では8本(!)の束にした電極をてんかん患者さんに埋め込んでいる。(本当に治療目的か??)
まあ、倫理面を別にすればかなりおもしろい論文。たとえば、Sydney Operaの建物の写真に反応する細胞は、「Sydney Opera」という文字にも同様に反応しており、pair-accosiate な情報をコードしていることがよく分かる。人間ならではの実験ね。
投稿者 sfujisawa : 18:10 | コメント (0)
JNS 6/22
Distinct Contributions of Hippocampal NMDA and AMPA Receptors to Encoding and Retrieval of One-Trial Place Memory
Tobias Bast, Bruno M. da Silva, and Richard G. M. Morris
Morrisのグループ。彼らの Nature 2003の焼き直し的論文かな。Task が One-Trial Place Memory taskに変わっているだけのような気がする
投稿者 sfujisawa : 18:08 | コメント (0)
2005年06月17日
Phys Rev Lett 6/17
Role of Delays in Shaping Spatiotemporal Dynamics of Neuronal Activity in Large Networks
Alex Roxin, Nicolas Brunel, and David Hansel
Alex Roxinはこの論文(PRL2004、Small-world neural networkでの同期形成のモデルの論文)の著者。Brunelは Amit や Wang XJ や Abbott とかとよく一緒に論文出している人ですね。
この論文は、networkにおいて、neuronどうしのinteractionのdelayを考慮すると、synchronizationのパターンに「phase」が存在するようになる、という内容。連続空間での firing rate modelと、Hodgikin-Huxley modelの、両方で計算している。
Communication Boundaries in Networks
A Trusina, M Rosvall, K Sneppen
「1960年代、スタンレー・ミルグラムという名のアメリカの心理学者は、人々をコミュニティに結びつけている複雑な人間関係の構図を捉えようとしていた。そのためにミルグラムは、カンザス州とネブラスカ州の住民から何人かをランダムに選び出して 160通の手紙を送りつけ、その手紙をボストンにいる彼の友人の株仲買人に転送してほしいと依頼した。ただし、友人の住所は知らせなかった。手紙を転送するにあたっては、それぞれの人の個人的な知り合いで、その株仲買人と社会的に「近い」と思われる人にだけ送るように頼んだ。最終的には、手紙の大半がボストンの友人のもとに届いたのである。手紙の多くは、6回前後の投函で着いたのである。ミルグラムの発見は、「6次の隔たり」(six degrees of separation) として人々に知られるようになった。どんな有名俳優とでも、6人の関係する人を通せば、結びついてしまうということになる。」(ここから引用)
この話はSmall-worldの例として有名な話よね。人間同士のネットワークは最短6人程度でつながっている、というのは良いとして、問題は、なぜ、手紙を送った人たちは、その最短に近い経路をたどることができたのか。
この論文ではそこに焦点を当てている。Real-world networkでのpath findingについて。
出発点 s から目的地 t に最短で到着できる確率 S(s→t)は、普通に式(1)で計算できる。これに加えて、自分の友達のうち、誰が多くのコネクションを持っているか、という情報を「betweenness」として重み付けに用いたときの確率 Sw(s→t)(式(2))を計算しているのがおもしろい。s→tまでの距離 l が小さいときと大きいときで、S(s→t)とSw(s→t)を計算している。その結果、real-world networkでは、距離 l が小さいときはS(s→t)が高く、距離 l が大きいときはSw(s→t)が高い、という結果が得られた。つまり、real-world networkで目的地までの最短のpathを探すには、pathのはじめの方ではbetweennessを重視して検索していき、目的地に近づいてきたら場当たり的に検索するのが良い、という結論。まあ、結論は当たり前っぽいけど、考え方と計算法がけっこうおもしろい論文でした。
投稿者 sfujisawa : 20:14 | コメント (0)
2005年06月16日
Current Issue
チェックだけ。
Neuron 6/16
Spatiotemporal Elements of Macaque V1 Receptive Fields
JNS 6/15
Functional Magnetic Resonance Imaging Activity during the Gradual Acquisition and Expression of Paired-Associate Memory
マカク猿のPaired-Associate Memoryで先駆的な研究をした宮下先生の論文をMethod以外全く引用していなかったから、pooneilさん怒っていますね。
投稿者 sfujisawa : 19:30 | コメント (0)
2005年06月09日
Nature 6/9
Uncovering the overlapping community structure of complex networks in nature and society
Complex networkのsubunit構造(彼らはこれを"comunity"と呼んでいる)に注目し、解析した論文。彼らの定義する"k-clique comunity"がよく分からなかったので内容が理解できませんでした。ごめんなさい。
投稿者 sfujisawa : 19:41 | コメント (0)
2005年06月03日
Phys Rev Lett 6/3
Synchronization is Enhanced in Weighted Complex Networks
タイトルにひかれて落としてみたけど、内容はよく分からん。どうやら考え方の枠組みは、L Pecoraさんという人の研究(これとかこれ)に端を発するみたい。ノード i の個々のダイナミクスが dx_i/dt = F(x_i)で表され、また他のノードへの出力をH(x_i)と表すとき、系のダイナミクスを dx_i/dt = F(x_i) + {Sigama}_j * G_ij * H(x_j) と考える枠組みらしい。正当っぽいけど、それだけにじっくり読まんとよく分からん。今回の論文は、どうも以前の論文にasynmetry性を加えただけのことらしい。
投稿者 sfujisawa : 18:33 | コメント (0)
2005年06月02日
Nature 6/2
Oxytocin increases trust in humans
「人を信頼させるホルモン確認」らしい。
投稿者 sfujisawa : 16:33 | コメント (0)
2005年06月01日
Neuron 6/2
The Hippocampal-VTA Loop: Controlling the Entry of Information into Long-Term Memory
Lisman JE と Grace AA という、ちょっと変わった組み合わせのReview。中脳の腹側被蓋野(VTA)のドーパミンニューロンは海馬CA1野に投射しているらしいのだが、これが、学習に関わるのかどうかを議論したReview。海馬とVTAがloopを形成していて、これが新規性の学習に機能的に働いている、というのが彼らの主張。あとでじっくり読んでみます。
The Neuroanatomy of Remote Memory
Squireのグループ。タイトルだけ見ると、Squireはいったい何をやったんだ、とびっくりしてしまいますが、 いつもの、側頭葉に傷害を持つ患者の記憶課題の実験でした。
実験は、側頭葉に障害を持つ患者と、側頭葉+別の皮質に傷害を持つ患者で、Autobiographic memoryの課題を行った。Autobiographic memoryは、傷害を受ける前の人生の、前1/3に起きたことを聞かれる記憶課題。側頭葉障害患者ではこの課題はできるが、側頭葉+別の皮質に障害を持つ患者では、この課題ができなかった、という結論。すなわち、長期記憶(Remote memory)は側頭葉だけに蓄えられるのではなく、皮質に広く蓄えられている可能性がある、という主張。
彼らの前報(Manns et al., neuron 2002 a,b)では、比較的短期(24hrとか)のpair-associate記憶課題は、海馬+側頭葉としている。また、Recall課題も、傷害の15年ぐらい前までのretrogradeの記憶は海馬+側頭葉としている。なので、今回用いたAutobiographic memoryは、単にエピソード記憶というだけでなく、Remote性がすごく強い課題である、ということを主張したいのだと思う。(しかし、彼らはなぜか自分たちの前報をひいていない。)
簡単にまとめると、人間では、比較的最近(ここ数年)のエピソード記憶は海馬+側頭葉に蓄えられ、すごい昔のエピソード記憶(remote memory)は、別の皮質に蓄えられると言うことかな。(Manns 2002 bのrecall課題をエピソード記憶と解釈すればの話しですが。)
投稿者 sfujisawa : 17:05 | コメント (0)
Nature Reviews Neuroscience 6月号
INTERACTIONS BETWEEN NUMBER AND SPACE IN PARIETAL CORTEX
おもしろそうだけどまだ読んでない。
投稿者 sfujisawa : 17:04 | コメント (0)
JNS 6/1
The Neural Correlates of Motor Skill Automaticity
投稿者 sfujisawa : 17:02 | コメント (0)
2005年05月25日
Nature Neurosci 6月号
Geometric and functional organization of cortical circuits
Svoboda研。光学系を駆使してcortexのmicrocircuitを調べる系の研究。手法的にも、内容的にも、Yoshimura&Dantzker&Callaway の Nature 2005 に似ていますね。
この論文では、somatosensory cortexのスライスを用いて、IV層・V層からII/III層へ錐体細胞がどのようなコネクションを形成しているのかをGeometry的(モルフォロジー的にシナプスを形成しているか)とfunction的(つまり、post-synaptic potential)に調べている。特に、barrel部とseptum部での違いを見ているところがおもしろい。
Dopaminergic modulation of limbic and cortical drive of nucleus accumbens in goal-directed behavior
Goto Y & Grace AA。Graceは久々に見た気がする。
Instructive signals for motor learning from visual cortical area MT
投稿者 sfujisawa : 17:52 | コメント (0)
JNS 5/25
チェックだけ。
Distinct Cortical Pathways for Processing Tool versus Animal Sounds
fMRI。
Functional Architecture of Spatial Attention in the Parietal Cortex of the Behaving Monkey
Synergy, Redundancy, and Independence in Population Codes, Revisited
Early Neural Correlates of Conscious Somatosensory Perception
投稿者 sfujisawa : 17:48 | コメント (0)
2005年05月24日
Learning&Memory 5月号
Recollection in an episodic-like memory task in the rat
Eacottら。Eacott & Norman JNS 2004 の続報的論文。
動物実験において、ある記憶課題が「Episode-like memory」課題であることを言うためには、what-where-whenそれぞれの情報をcueとして含む課題である必要があると言われている。しかし、「when」情報をcueとする課題は難しく、いままでrodentの実験ではEpisode-like memory課題はまだ構築されていない。
Eacott JNS 2004 では、「when」情報の代わりに「context」情報を用いて、what-where-contextのcueを必要とする系を立ち上げていた。ただし、課題としてobject recognition taskを用いているため、「familiarity」性の強い記憶課題であった。(Episode-like memoryは「recollection」性の強い記憶だと考えられるので、そこが彼らは不満だったのでしょう。)
今回は、改良を加えることにより、より「recollection」性の強い系にした、という論文。具体的には、objectを同時には見えない位置に置くことにより、recollectionが必要な課題にしたという内容。(taskは object recognition taskのまま。)この課題が海馬依存であるかどうかまでは調べていない。
Acetylcholine in the orbitofrontal cortex is necessary for the acquisition of a socially transmitted food preference
Eichenbaumのグループ。
投稿者 sfujisawa : 17:56 | コメント (0)
RevModPhys (Current Issue)
The Kuramoto model: A simple paradigm for synchronization phenomena
蔵本モデルのReview。ニューラルネットやHopfieldモデルとの関連についてもちょろっと書いてある。
投稿者 sfujisawa : 11:56 | コメント (0)
2005年05月18日
Nature 5/19
Sustained firing in auditory cortex evoked by preferred stimuli. マカクザルのauditory cortexでの、stimulus evoked な persistent activityについて。auditory cortexのneuronでは、ある時間音刺激を与えた場合でもはじめのある期間しか発火しないらしいのだが、preferな音刺激な場合では、音刺激を与えている期間中、発火し続けるというもの。
Lack of long-term cortical reorganization after macaque retinal lesions. Logothetisのグループ。(難しそうなのでちょろっと目を通した程度。)マカクザルの網膜をlesionしたとき、V1野ではネットワークの再編が起こる、と今までは言われていたが、実はそうでもない、ということを示した論文。fMRIを用いてV1野の活動を広域に記録し、かつ電気生理で確認もしている。fMRIでここまでできるんだね。
投稿者 sfujisawa : 19:24 | コメント (0)
JNS 5/18
Variation of Input-Output Properties along the Somatodendritic Axis of Pyramidal Neurons. Oviedo H & Reyes AD。Reyesは、最近はネットワークでのinput-output property(とくにsynfireとか)をやっていたけど、今回は、dendrite演算系をやっていますね。
Multisensory Integration of Dynamic Faces and Voices in Rhesus Monkey Auditory Cortex. Logothetisのグループ。
投稿者 sfujisawa : 19:20 | コメント (0)
2005年05月13日
Nature 5/12
The origin of bursts and heavy tails in human dynamics. Barabasi。せっかく論文が読める環境になった、と思ったら、Nature 落ちてる。。
投稿者 sfujisawa : 15:56 | コメント (0)
2005年05月08日
Neuron 5/3
Fast Odor Learning Improves Reliability of Odor Responses in the Locust Antennal Lobe Laurent のグループ
Associative Learning Engages the Hippocampus and Interacts with Explicit Associative Learning
投稿者 sfujisawa : 22:34 | コメント (0)
2005年05月06日
Science 5/6
Target Cell-Dependent Normalization of Transmitter Release at Neocortical Synapses Daniel Johnstonのグループ。
Attractor Dynamics in the Hippocampal Representation of the Local Environment
O'Keefe のグループ。SFN2004で発表していたやつ。Place cell は、異なった空間(ここでは丸い空間と四角い空間)において発火する領域(place field)が異なる。では、丸い空間を徐々に四角い空間に変えていったら、place cell はどのように発火するのだろうか、という実験。ここでは、丸い空間を徐々に四角い空間に変えていったら、place cell の place field は、ある時点で、不連続に、丸い空間の place field から四角い空間の place field へ遷移するという結果が得られている。つまり、丸と四角の中間の空間であっても、丸い空間か四角い空間のどちらかに認識されているわけで、O'Keefe たちは、これを place cell の(あるいは place cell assembly の)"attractor"である、としている。
O'Keefe Nature 2002 の続き的な論文かな?Nature 2002 は、Place cell が異なった空間において異なった place field を構成するまでの過程を追った論文で、これもすばらしい論文。
投稿者 sfujisawa : 20:57 | コメント (0)
2005年05月02日
PNAS Identification of network-level coding units for real-time representation of episodic experiences in the hippocampus. JZ Tsein のグループ
マウスでも使える小型の96channelの電極を開発した、という論文。260個のneuronの発火を分離できるとのこと。ただ、課題は3種類のstartle刺激(エアパフなど)を与えただけのもので、「episodic」とタイトルにいれるのはちとまずいかと。
